

Review
doi: 10.1016/j.semcdb.2010.02.004.
Epub 2010 Feb 10.
MicroRNAs in mouse development and disease
Affiliations
- PMID: 20152923
- PMCID: PMC2938480
- DOI: 10.1016/j.semcdb.2010.02.004
Item in Clipboard
Review
MicroRNAs in mouse development and disease
Semin Cell Dev Biol.
2010 Sep.
Abstract
MicroRNAs, small non-coding RNAs which act as repressors of target genes, were discovered in 1993, and since then have been shown to play important roles in the development of numerous systems. Consistent with this role, they are also implicated in the pathogenesis of multiple diseases. Here we review the involvement of microRNAs in mouse development and disease, with particular reference to deafness as an example.
(c) 2010 Elsevier Ltd. All rights reserved.
Figures

Cartoon showing putative mechanisms of microRNA repression. The microRNA is represented by a thick red block over the violet line, which is the target mRNA. The blue hexagon represents the translation initiation complex and the orange circles ribosomes. (A) Perfect binding of miRNA to target mRNA and cleavage of mRNA followed by degradation. (B) Inhibition of translation initiation. (C) Inhibition of elongation/termination and promotion of ribosome drop-off. (D) Recruitment of a proteolytic enzyme to degrade emerging polypeptides. (E) Deadenylation, followed by degradation, of the mRNA .

Scanning electron microscopy of diminuendo homozygote and heterozygote hair cells at 5 days old. (A–C) Outer hair cells of wildtype (A), heterozygote (B) and homozygote (C) mice at 5 days old. Heterozygote hair cells appear indistinguishable from wildtype hair cells at this early stage, but homozygote hair cells show ectopic stereocilia and disorganisation. Scale bars = 5 μm.
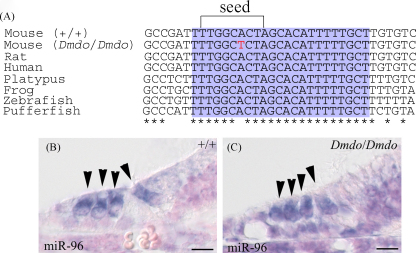
miR-96 conservation and expression. (A) Alignment of DNA sequences from wildtype mouse, diminuendo homozygote, rat, human, platypus, frog, zebrafish and pufferfish. The purple region shows the mature miRNA sequence for each species, and the bracket delineates the seed region critical for target binding. The mutation in the seed region is indicated by the red letter. The mature sequence is absolutely conserved between the species shown, and also between cow, dog, horse, macaque, opossum, chimpanzee, orang-utan, ground squirrel, tree shrew, mouse lemur, bushbaby, cat, armadillo, tenrec, medaka, rabbit, stickleback and tetraodon (sequences obtained from Ensembl v50; http://www.ensembl.org ). (B and C) Expression of Mir96 at P5 in wildtype (B) and homozygote (C). Hair cells are marked by arrowheads. Scale bars = 10 μm. Part A reprinted with permission from Lewis et al. .

Hypergeometric analysis of microarray data comparing diminuendo homozygote to wildtype at 4 days old. (A) Microarray analysis showing enrichment and depletion of heptamers in 3′UTRs using Sylamer. The x-axis represents the sorted gene list from most upregulated (left) to most downregulated (right). The y-axis shows the hypergeometric significance for enrichment or depletion of heptamers in 3′UTRs of the genes under consideration when compared to the 3′UTRs in the complementary gene set. Positive values indicate enrichment (−log10(P-value)) and negative values depletion (log10(P-value)). Any heptamers which are present more often than would be expected will stand out above the significance line (adjusted P-value = 0.01), and any which are present less often than would be expected will stand out below the significance line on the negative axis. (B) The same analysis as in A, where each 3′UTR has been replaced by the concatenation of its orthologous 3′UTRs in human and rat. The seed match for the wildtype miR-96 shows similar enrichment as compared with the analysis in A. In contrast, the enrichment of the miR-96 diminuendo mutant binding sites in the downregulated genes is barely above background (dotted line). Reprinted with permission from Lewis et al. .

Ocm, Slc26a5, Gfi1, Ptprq and Pitpnm1 in diminuendo. (A) Quantitative real-time PCR on cDNA generated from normalised RNA from the organs of Corti of 4-day-old littermates. Ocm, Slc26a5 (prestin), Ptprq, Pitpnm1 and Gfi1 are downregulated in homozygotes. Error bars represent standard deviation. Quantities normalised to Hprt1 levels; Ngfr is expressed in support cells adjacent to hair cells and was used to assess the quantity of sensory material. Three animals were used per genotype and DNA from each was run in triplicate. (B–E) Location of oncomodulin (B and C) and prestin (D and E) in 5-day-old wildtype (B and D) and homozygote (C and E) littermates. Scale bars = 10 μm.
References
-
- Chalfie M., Horvitz H.R., Sulston J.E. Mutations that lead to reiterations in the cell lineages of C. elegans. Cell. 1981;24:59–69. - PubMed
-
- Lee R.C., Feinbaum R.L., Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854. - PubMed
-
- Reinhart B.J., Slack F.J., Basson M., Pasquinelli A.E., Bettinger J.C., Rougvie A.E. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. 2000;403:901–906. - PubMed
-
- Pasquinelli A.E., Reinhart B.J., Slack F., Martindale M.Q., Kuroda M.I., Maller B. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature. 2000;408:86–89. - PubMed
-
- Molnár A., Schwach F., Studholme D.J., Thuenemann E.C., Baulcombe D.C. miRNAs control gene expression in the single-cell alga Chlamydomonas reinhardtii. Nature. 2007;447:1126–1129. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
