

Learning to see: patterned visual activity and the development of visual function
- PMID: 20153060
- PMCID: PMC3415282
- DOI: 10.1016/j.tins.2010.01.003
Learning to see: patterned visual activity and the development of visual function
Abstract
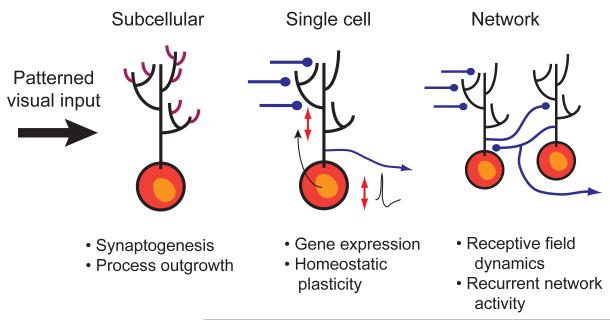
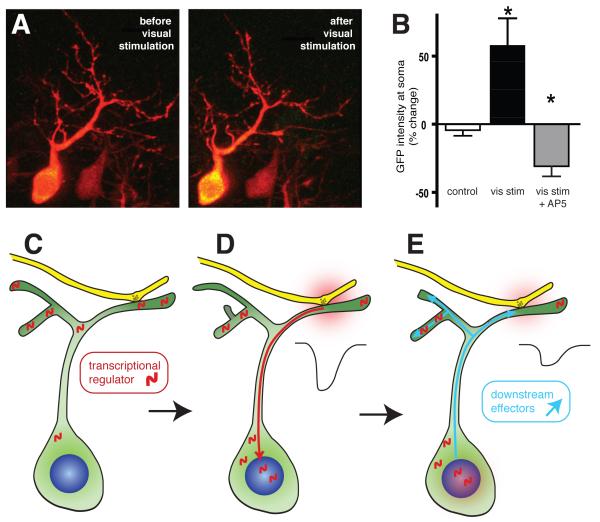
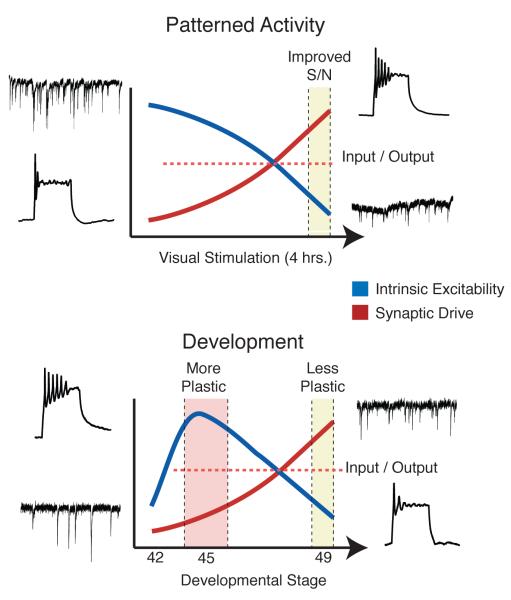
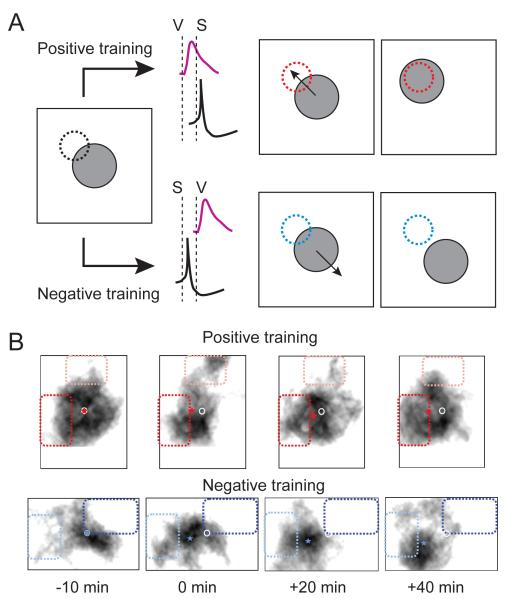
To successfully interact with their environments, developing organisms need to correctly process sensory information and generate motor outputs appropriate to their size and structure. Patterned sensory experience has long been known to induce various forms of developmental plasticity that ultimately shape mature neural circuits. These same types of plasticity also allow developing organisms to respond appropriately to the external world by dynamically adapting neural circuit function to ongoing changes in brain circuitry and sensory input. Recent work on the visual systems of frogs and fish has provided an unprecedented view into how visual experience dynamically affects circuit function at many levels, ranging from gene expression to network function, ultimately leading to system-wide functional adaptations.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Mouse vision as a gateway for understanding how experience shapes neural circuits.Front Neural Circuits. 2014 Oct 2;8:123. doi: 10.3389/fncir.2014.00123. eCollection 2014. Front Neural Circuits. 2014. PMID: 25324730 Free PMC article. Review.
-
Rapid Sensory Adaptation Redux: A Circuit Perspective.Neuron. 2016 Oct 19;92(2):298-315. doi: 10.1016/j.neuron.2016.09.046. Neuron. 2016. PMID: 27764664 Free PMC article. Review.
-
Visual activity and cortical rewiring: activity-dependent plasticity of cortical networks.Prog Brain Res. 2006;157:3-11. doi: 10.1016/s0079-6123(06)57001-3. Prog Brain Res. 2006. PMID: 17167899 Review.
-
Visual circuits in flies: beginning to see the whole picture.Curr Opin Neurobiol. 2015 Oct;34:125-32. doi: 10.1016/j.conb.2015.03.010. Epub 2015 Apr 14. Curr Opin Neurobiol. 2015. PMID: 25881091 Free PMC article. Review.
-
Nogo Receptor 1 Confines a Disinhibitory Microcircuit to the Critical Period in Visual Cortex.J Neurosci. 2016 Oct 26;36(43):11006-11012. doi: 10.1523/JNEUROSCI.0935-16.2016. J Neurosci. 2016. PMID: 27798181 Free PMC article.
Cited by
-
Epigenetic regulation of GABAergic differentiation in the developing brain.Front Cell Neurosci. 2022 Sep 23;16:988732. doi: 10.3389/fncel.2022.988732. eCollection 2022. Front Cell Neurosci. 2022. PMID: 36212693 Free PMC article.
-
GABA expression and regulation by sensory experience in the developing visual system.PLoS One. 2012;7(1):e29086. doi: 10.1371/journal.pone.0029086. Epub 2012 Jan 5. PLoS One. 2012. PMID: 22242157 Free PMC article.
-
Heterogeneity and dynamics of lateral line afferent innervation during development in zebrafish (Danio rerio).J Comp Neurol. 2012 May 1;520(7):1376-86. doi: 10.1002/cne.22798. J Comp Neurol. 2012. PMID: 22102005 Free PMC article.
-
Visual experience-dependent maturation of correlated neuronal activity patterns in a developing visual system.J Neurosci. 2011 Jun 1;31(22):8025-36. doi: 10.1523/JNEUROSCI.5802-10.2011. J Neurosci. 2011. PMID: 21632924 Free PMC article.
-
Synapse maturation is enhanced in the binocular region of the retinocollicular map prior to eye opening.J Neurophysiol. 2012 Jun;107(11):3200-16. doi: 10.1152/jn.00943.2011. Epub 2012 Mar 7. J Neurophysiol. 2012. PMID: 22402661 Free PMC article.
References
-
- Ruthazer ES. You’re perfect, now change--redefining the role of developmental plasticity. Neuron. 2005;45:825–828. - PubMed
-
- Katz LC, Shatz CJ. Synaptic activity and the construction of cortical circuits. Science. 1996;274:1133–1138. - PubMed
-
- Wiesel TN, Hubel DH. Single-Cell Responses in Striate Cortex of Kittens Deprived of Vision in One Eye. J Neurophysiol. 1963;26:1003–1017. - PubMed
-
- Hooks BM, Chen C. Distinct roles for spontaneous and visual activity in remodeling of the retinogeniculate synapse. Neuron. 2006;52:281–291. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
