

Paramutation in maize: RNA mediated trans-generational gene silencing
- PMID: 20153628
- PMCID: PMC2859986
- DOI: 10.1016/j.gde.2010.01.008
Paramutation in maize: RNA mediated trans-generational gene silencing
Abstract
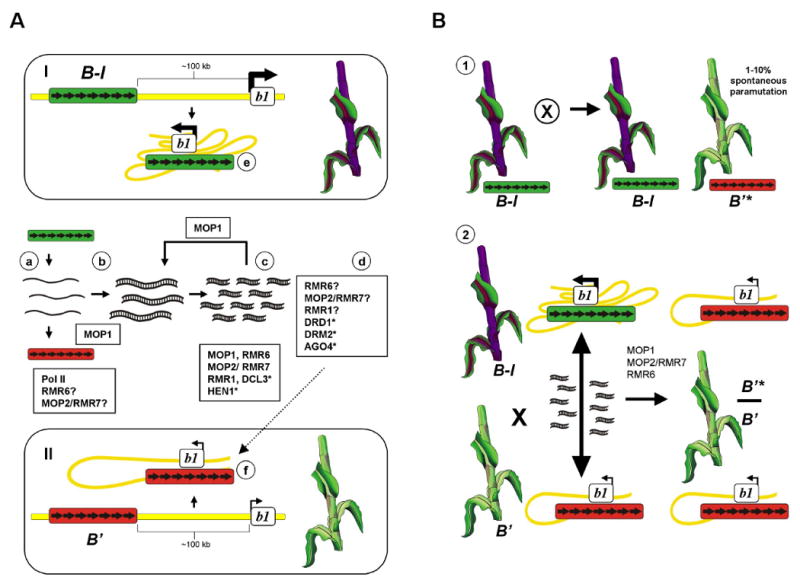
Paramutation involves trans-interactions between alleles or homologous sequences that establish distinct gene expression states that are heritable for generations. It was first described in maize by Alexander Brink in the 1950s, with his studies of the red1 (r1) locus. Since that time, paramutation-like phenomena have been reported in other maize genes, other plants, fungi, and animals. Paramutation can occur between endogenous genes, two transgenes or an endogenous gene, and transgene. Recent results indicate that paramutation involves RNA-mediated heritable chromatin changes and a number of genes implicated in RNAi pathways. However, not all aspects of paramutation can be explained by known mechanisms of RNAi-mediated transcriptional silencing.
2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Pilu R, Panzeri D, Cassani E, Cerino Badone F, Landoni M, Nielsen E. A paramutation phenomenon is involved in the genetics of maize low phytic acid1-241 (lpa1-241) trait. Heredity. 2009;102:236–245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
