

Review
doi: 10.1016/j.gde.2010.01.003.
Epub 2010 Feb 12.
New functions for an old variant: no substitute for histone H3.3
Affiliations
- PMID: 20153629
- PMCID: PMC2860041
- DOI: 10.1016/j.gde.2010.01.003
Item in Clipboard
Review
New functions for an old variant: no substitute for histone H3.3
Curr Opin Genet Dev.
2010 Apr.
Abstract
Histone proteins often come in different variants serving specialized functions in addition to their fundamental role in packaging DNA. The metazoan histone H3.3 has been most closely associated with active transcription. Its role in histone replacement at active genes and promoters is conserved to the single histone H3 in yeast. However, recent genetic studies in flies have challenged its importance as a mark of active chromatin, and revealed unexpected insights into essential functions of H3.3 in the germline. With strikingly little amino acid sequence difference to the canonical H3, H3.3 therefore accomplishes a surprising variety of cellular and developmental processes.
2010 Elsevier Ltd. All rights reserved.
Figures
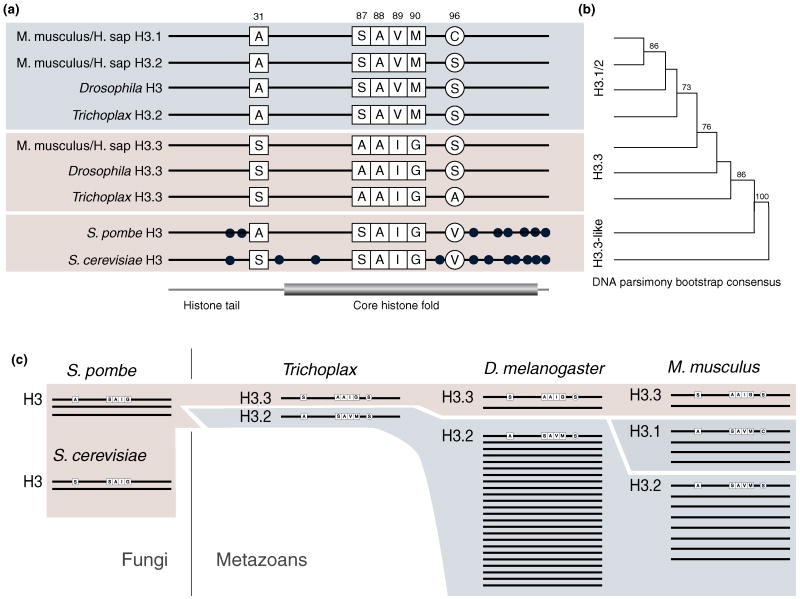
(a) Schematic representation of the major non-centromeric histone H3 protein sequences from human, mouse and Trichoplax adhearens (one of the most basal metazoan species), as well as budding and fission yeast. Amino acids that distinguish variants are highlighted with residue numbers, additional differences are indicated as dots. H3.1 only exists in mammals and only differs in position 96 from H3.2 present in all metazoans. H3.2 and H3.3 are distinguished by one amino acid difference at position 31 in the histone tail and three in amino acids 87-90 in the core histone fold. (b) Phylogenetic relationship of the respective histone H3 genes. An unrooted parsimony tree was constructed based on representative coding sequences (consensus tree of 100 bootstraps, excluding the wobble bases). The H3 genes of S. pombe and S. cerevisiae cluster with metazoan H3.3. (c) Schematic overview of the major non-centromeric gene complements of the indicated species. The placozoan Trichoplax adhearens has only one gene for H3.2 and H3.3 each, while higher metazoans have greatly expanded H3.1/2 gene complements.
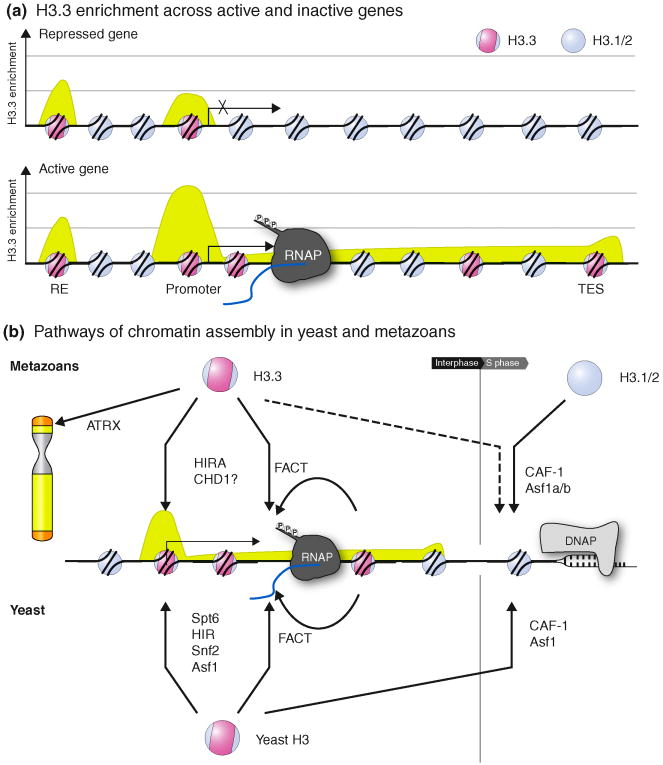
(a) Schematic map of an active and inactive gene locus comprising an upstream regulatory element (RE), transcription start site (Promoter) and transcription end site (TES). The distribution of histone H3.3 across the locus is shown in green, with representative H3.3 and H3.1/2 nucleosomes, as well as RNA polymerase II (RNAP). (b) Summary of the known factors involved in replication-dependent (right, S phase) and replication-independent (left, Interphase) chromatin assembly pathways, in metazoans and yeast. Replication-coupled assembly is thought to be mediated by the CAF-1 complex and Asf1 proteins in the wake of DNA polymerase (DNAP). H3.3-enrichment at telomeres is dependent on ATRX. Replication-independent deposition at promoters, regulatory elements and genic regions in metazoans requires HIRA, CHD1, and/or other factors, analogous to pathways in yeast mediated by Snf2, Asf1, HIR complex and/or Spt6. The FACT complex (Spt16 and Pob3/Ssrp1) might contribute to incorporation of new or recycling of old histones.
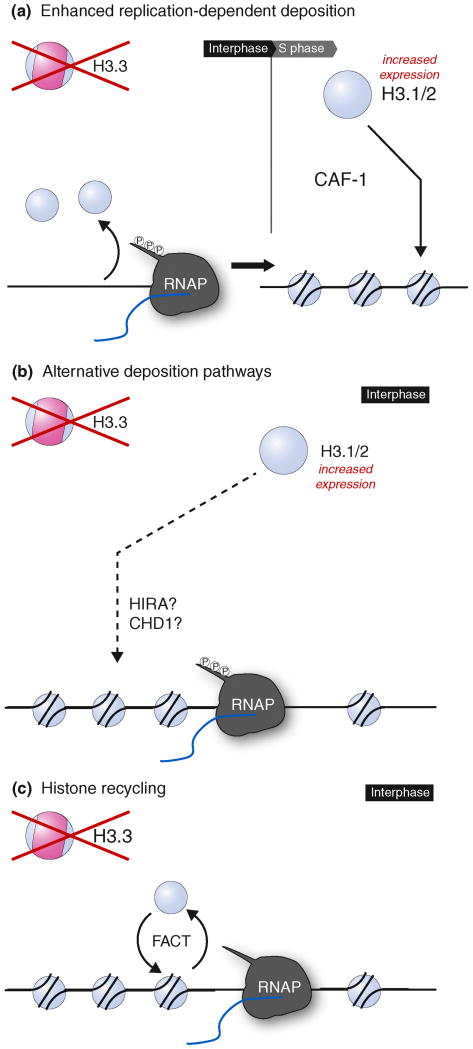
(a) Absence of H3.3 as substrate for replication-independent chromatin assembly could create nucleosome-free regions (see also ref. [62]). In rapidly dividing cells, these gaps could be filled during the next S phase via canonical replication-dependent chromatin assembly. (b) Elevated levels of H3.1/2 throughout the cell cycle could provide substrate for replication-independent chromatin assembly factors that are not restricted to H3.3. (c) In the absence of de-novo chromatin assembly, the FACT complex could favor transient eviction and redeposition of histone units in cis.
References
-
- Franklin SG, Zweidler A. Non-allelic variants of histones 2a, 2b and 3 in mammals. Nature. 1977;266:273–275. - PubMed
-
- Ahmad K, Henikoff S. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol Cell. 2002;9:1191–1200. - PubMed
-
- Marzluff WF, Gongidi P, Woods KR, Jin J, Maltais LJ. The human and mouse replication-dependent histone genes. Genomics. 2002;80:487–498. - PubMed
-
- Rooney AP, Piontkivska H, Nei M. Molecular evolution of the nontandemly repeated genes of the histone 3 multigene family. Molecular Biology and Evolution. 2002;19:68–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
