

Protecting against antimicrobial effectors in the phagosome allows SodCII to contribute to virulence in Salmonella enterica serovar Typhimurium
- PMID: 20154132
- PMCID: PMC2849443
- DOI: 10.1128/JB.00016-10
Protecting against antimicrobial effectors in the phagosome allows SodCII to contribute to virulence in Salmonella enterica serovar Typhimurium
Abstract
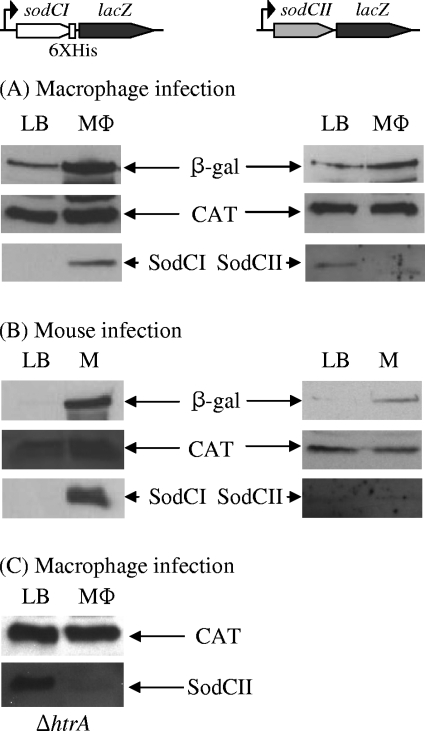
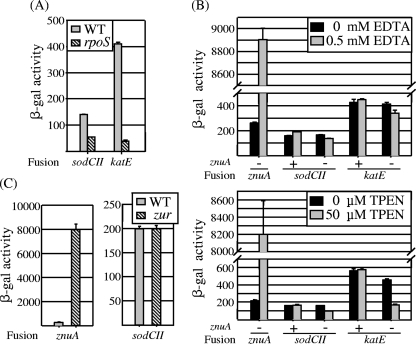
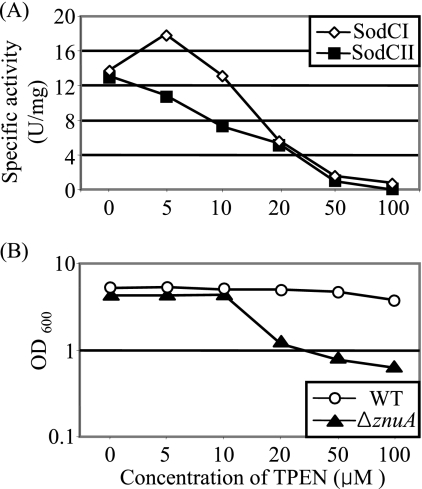
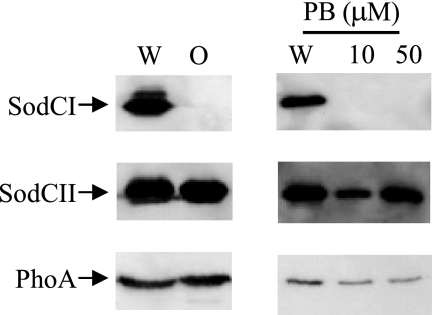
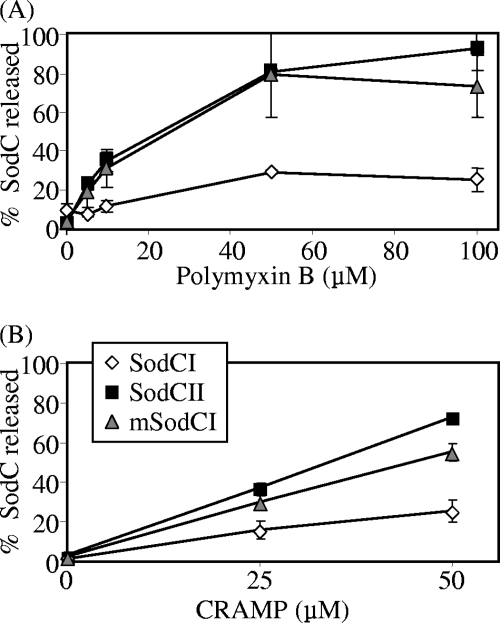
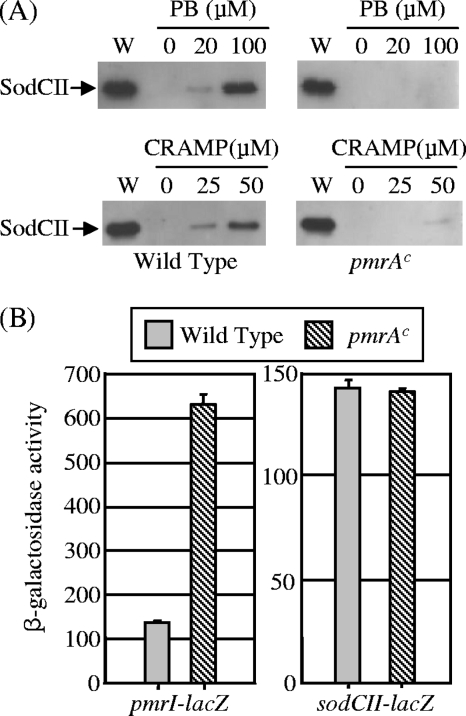
Salmonella enterica serovar Typhimurium replicates in macrophages, where it is subjected to antimicrobial substances, including superoxide, antimicrobial peptides, and proteases. The bacterium produces two periplasmic superoxide dismutases, SodCI and SodCII. Although both are expressed during infection, only SodCI contributes to virulence in the mouse by combating phagocytic superoxide. The differential contribution to virulence is at least partially due to inherent differences in the SodCI and SodCII proteins that are independent of enzymatic activity. SodCII is protease sensitive, and like other periplasmic proteins, it is released by osmotic shock. In contrast, SodCI is protease resistant and is retained within the periplasm after osmotic shock, a phenomenon that we term "tethering." We hypothesize that in the macrophage, antimicrobial peptides transiently disrupt the outer membrane. SodCII is released and/or phagocytic proteases gain access to the periplasm, and SodCII is degraded. SodCI is tethered within the periplasm and is protease resistant, thereby remaining to combat superoxide. Here we test aspects of this model. SodCII was released by the antimicrobial peptide polymyxin B or a mouse macrophage antimicrobial peptide (CRAMP), while SodCI remained tethered within the periplasm. A Salmonella pmrA constitutive mutant no longer released SodCII in vitro. Moreover, in the constitutive pmrA background, SodCII could contribute to survival of Salmonella during infection. SodCII also provided a virulence benefit in mice genetically defective in production of CRAMP. Thus, consistent with our model, protecting the outer membrane against antimicrobial peptides allows SodCII to contribute to virulence in vivo. These data also suggest direct in vivo cooperative interactions between macrophage antimicrobial effectors.
Figures
References
-
- Ammendola, S., M. Ajello, P. Pasquali, J. S. Kroll, P. R. Langford, G. Rotilio, P. Valenti, and A. Battistoni. 2005. Differential contribution of SodC1 and SodC2 to intracellular survival and pathogenicity of Salmonella enterica serovar Choleraesuis. Microbes Infect. 7:698-707. - PubMed
-
- Ammendola, S., P. Pasquali, F. Pacello, G. Rotilio, M. Castor, S. J. Libby, N. Figueroa-Bossi, L. Bossi, F. C. Fang, and A. Battistoni. 2008. Regulatory and structural differences in the Cu,Zn-superoxide dismutases of Salmonella enterica and their significance for virulence. J. Biol. Chem. 283:13688-13699. - PMC - PubMed
-
- Bader, M. W., S. Sanowar, M. E. Daley, A. R. Schneider, U. Cho, W. Xu, R. E. Klevit, M. H. Le, and S. I. Miller. 2005. Recognition of antimicrobial peptides by a bacterial sensor kinase. Cell 122:461-472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
