

Characterizing the dynamics of CD4+ T cell priming within a lymph node
- PMID: 20154206
- PMCID: PMC3153313
- DOI: 10.4049/jimmunol.0903117
Characterizing the dynamics of CD4+ T cell priming within a lymph node
Abstract
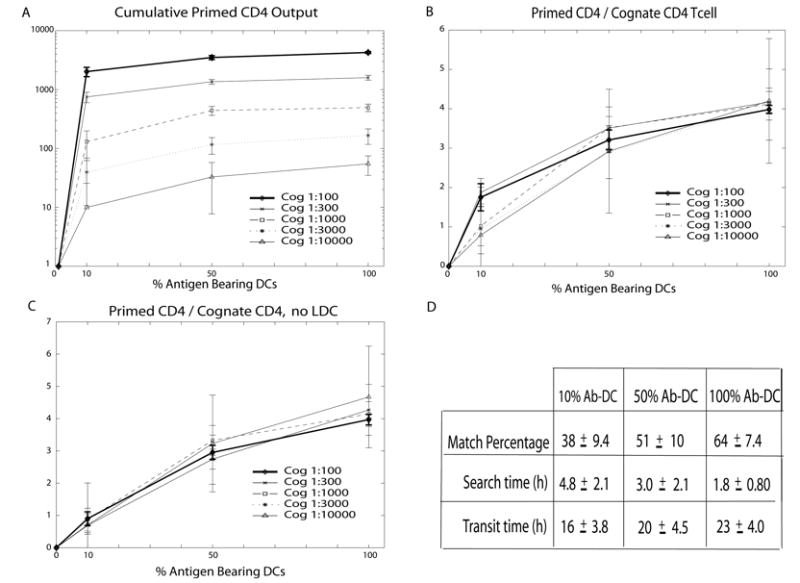
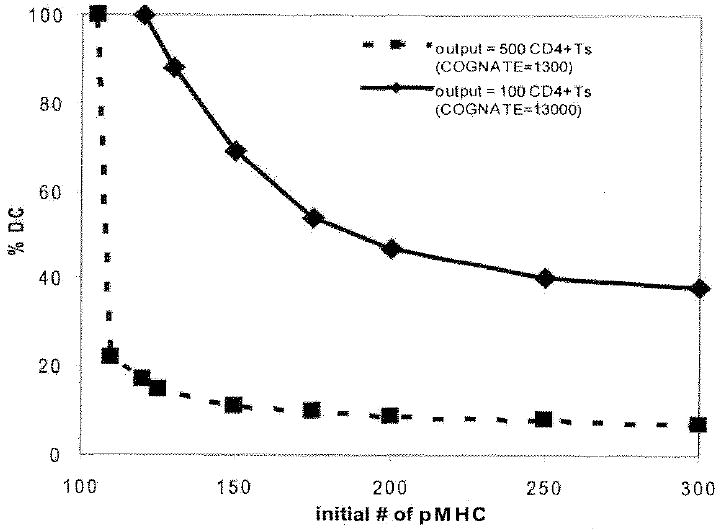
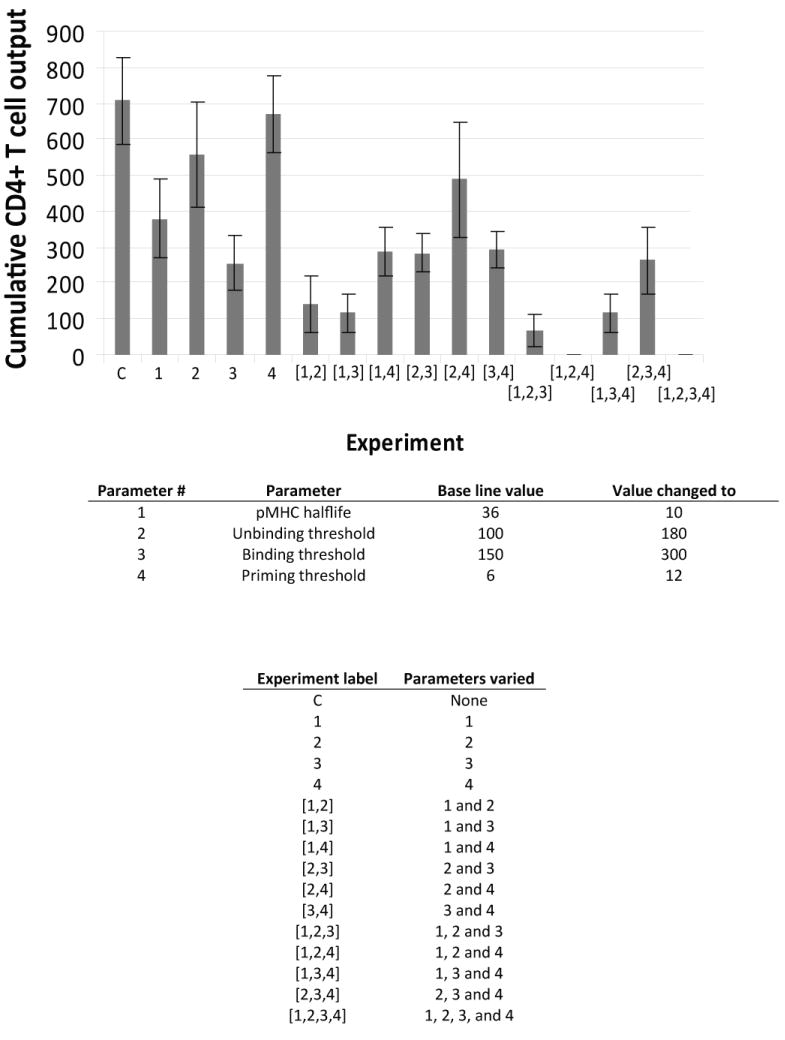
Generating adaptive immunity postinfection or immunization requires physical interaction within a lymph node T zone between Ag-bearing dendritic cells (DCs) and rare cognate T cells. Many fundamental questions remain regarding the dynamics of DC-CD4+ T cell interactions leading to priming. For example, it is not known how the production of primed CD4+ T cells relates to the numbers of cognate T cells, Ag-bearing DCs, or peptide-MHCII level on the DC. To address these questions, we developed an agent-based model of a lymph node to examine the relationships among cognate T cell frequency, DC density, parameters characterizing DC-T cell interactions, and the output of primed T cells. We found that the output of primed CD4+ T cells is linearly related to cognate frequency, but nonlinearly related to the number of Ag-bearing DCs present during infection. This addresses the applicability of two photon microscopy studies to understanding actual infection dynamics, because these types of experiments increase the cognate frequency by orders of magnitude compared with physiologic levels. We found a trade-off between the quantity of peptide-major histocompatibility class II on the surface of individual DCs and number of Ag-bearing DCs present in the lymph node in contributing to the production of primed CD4+ T cells. Interestingly, peptide-major histocompatibility class II t(1/2) plays a minor, although still significant, role in determining CD4+ T cell priming, unlike the primary role that has been suggested for CD8+ T cell priming. Finally, we identify several pathogen-targeted mechanisms that, if altered in their efficiency, can significantly effect the generation of primed CD4+ T cells.
Figures
Similar articles
-
Predicting lymph node output efficiency using systems biology.J Theor Biol. 2013 Oct 21;335:169-84. doi: 10.1016/j.jtbi.2013.06.016. Epub 2013 Jun 29. J Theor Biol. 2013. PMID: 23816876 Free PMC article.
-
Spatiotemporally Distinct Interactions with Dendritic Cell Subsets Facilitates CD4+ and CD8+ T Cell Activation to Localized Viral Infection.Immunity. 2015 Sep 15;43(3):554-65. doi: 10.1016/j.immuni.2015.07.020. Epub 2015 Aug 18. Immunity. 2015. PMID: 26297566
-
A comparison of random vs. chemotaxis-driven contacts of T cells with dendritic cells during repertoire scanning.J Theor Biol. 2008 Feb 21;250(4):732-51. doi: 10.1016/j.jtbi.2007.10.015. Epub 2007 Oct 18. J Theor Biol. 2008. PMID: 18068193 Free PMC article.
-
Studying interactions between dendritic cells and T cells in vivo.Curr Opin Immunol. 2019 Jun;58:24-30. doi: 10.1016/j.coi.2019.02.002. Epub 2019 Mar 15. Curr Opin Immunol. 2019. PMID: 30884422 Free PMC article. Review.
-
T-cell activation by dendritic cells in the lymph node: lessons from the movies.Nat Rev Immunol. 2008 Sep;8(9):675-84. doi: 10.1038/nri2379. Nat Rev Immunol. 2008. PMID: 19172690 Review.
Cited by
-
Agent-based model predicts that layered structure and 3D movement work synergistically to reduce bacterial load in 3D in vitro models of tuberculosis granuloma.PLoS Comput Biol. 2024 Jul 12;20(7):e1012266. doi: 10.1371/journal.pcbi.1012266. eCollection 2024 Jul. PLoS Comput Biol. 2024. PMID: 38995971 Free PMC article.
-
Phenotypic transition maps of 3D breast acini obtained by imaging-guided agent-based modeling.Integr Biol (Camb). 2011 Apr;3(4):408-21. doi: 10.1039/c0ib00092b. Epub 2011 Mar 4. Integr Biol (Camb). 2011. PMID: 21373705 Free PMC article.
-
Defining the quantitative limits of intravital two-photon lymphocyte tracking.Proc Natl Acad Sci U S A. 2011 Jul 26;108(30):12401-6. doi: 10.1073/pnas.1102288108. Epub 2011 Jul 6. Proc Natl Acad Sci U S A. 2011. PMID: 21734152 Free PMC article.
-
A brief overview of mathematical modeling of the within-host dynamics of Mycobacterium tuberculosis.Front Appl Math Stat. 2024;10:1355373. doi: 10.3389/fams.2024.1355373. Epub 2024 Jan 30. Front Appl Math Stat. 2024. PMID: 39906541 Free PMC article.
-
Agent-based models of inflammation in translational systems biology: A decade later.Wiley Interdiscip Rev Syst Biol Med. 2019 Nov;11(6):e1460. doi: 10.1002/wsbm.1460. Epub 2019 Jul 1. Wiley Interdiscip Rev Syst Biol Med. 2019. PMID: 31260168 Free PMC article. Review.
References
-
- Randolph GJ, Angeli V, Swartz MA. Dendritic-cell trafficking to lymph nodes through lymphatic vessels. Nat Rev Immunol. 2005;5:617–628. - PubMed
-
- von Andrian UH, Mempel TR. Homing and cellular traffic in lymph nodes. Nat Rev Immunol. 2003;3:867–878. - PubMed
-
- Catron DM, Itano AA, Pape KA, Mueller DL, Jenkins MK. Visualizing the first 50 hr of the primary immune response to a soluble antigen. Immunity. 2004;21:341–347. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
