

An ovine transgenic Huntington's disease model
- PMID: 20154343
- PMCID: PMC2860888
- DOI: 10.1093/hmg/ddq063
An ovine transgenic Huntington's disease model
Abstract
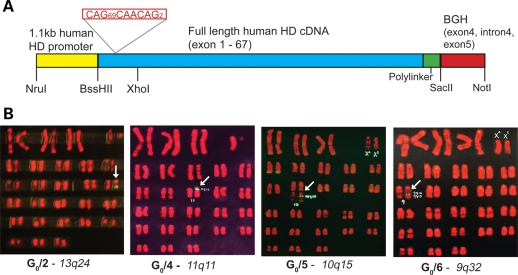
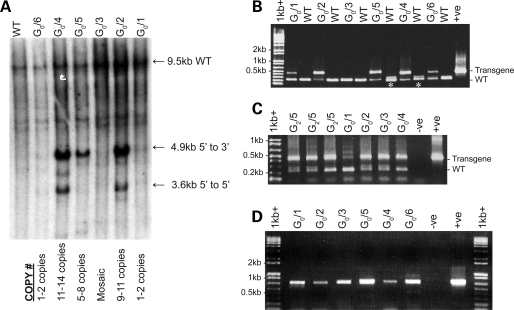
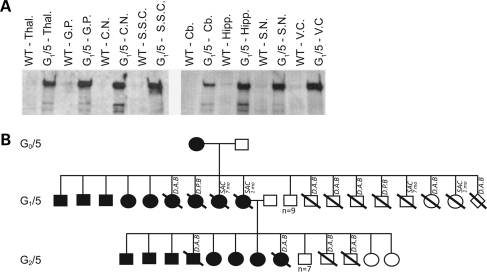
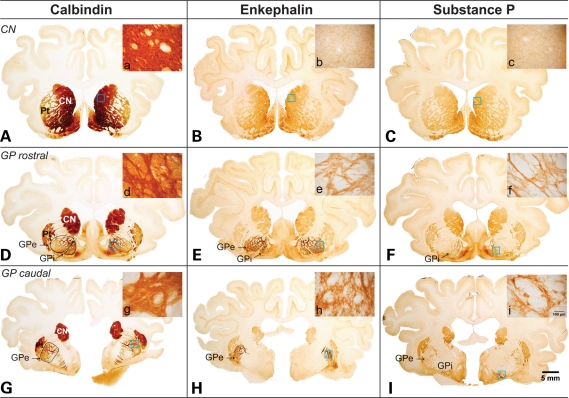
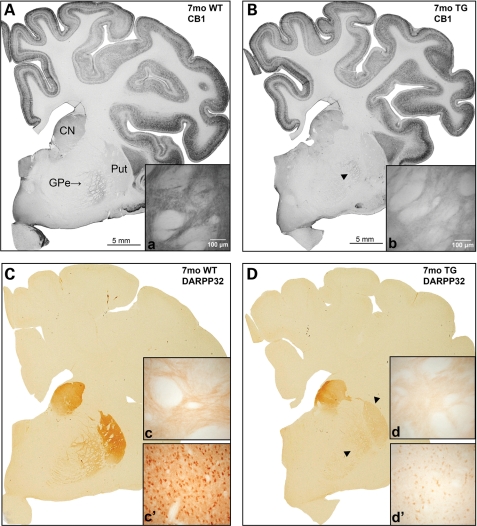
Huntington's disease (HD) is an inherited autosomal dominant neurodegenerative disorder caused by an expansion of a CAG trinucleotide repeat in the huntingtin (HTT) gene [Huntington's Disease Collaborative Research Group (1993) A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. The Huntington's Disease Collaborative Research Group. Cell, 72, 971-983]. Despite identification of the gene in 1993, the underlying life-long disease process and effective treatments to prevent or delay it remain elusive. In an effort to fast-track treatment strategies for HD into clinical trials, we have developed a new large-animal HD transgenic ovine model. Sheep, Ovis aries L., were selected because the developmental pattern of the ovine basal ganglia and cortex (the regions primarily affected in HD) is similar to the analogous regions of the human brain. Microinjection of a full-length human HTT cDNA containing 73 polyglutamine repeats under the control of the human promotor resulted in six transgenic founders varying in copy number of the transgene. Analysis of offspring (at 1 and 7 months of age) from one of the founders showed robust expression of the full-length human HTT protein in both CNS and non-CNS tissue. Further, preliminary immunohistochemical analysis demonstrated the organization of the caudate nucleus and putamen and revealed decreased expression of medium size spiny neuron marker DARPP-32 at 7 months of age. It is anticipated that this novel transgenic animal will represent a practical model for drug/clinical trials and surgical interventions especially aimed at delaying or preventing HD initiation. New sequence accession number for ovine HTT mRNA: FJ457100.
Figures
Similar articles
-
Transgenic mice expressing mutated full-length HD cDNA: a paradigm for locomotor changes and selective neuronal loss in Huntington's disease.Philos Trans R Soc Lond B Biol Sci. 1999 Jun 29;354(1386):1035-45. doi: 10.1098/rstb.1999.0456. Philos Trans R Soc Lond B Biol Sci. 1999. PMID: 10434303 Free PMC article.
-
Towards a transgenic model of Huntington's disease in a non-human primate.Nature. 2008 Jun 12;453(7197):921-4. doi: 10.1038/nature06975. Epub 2008 May 18. Nature. 2008. PMID: 18488016 Free PMC article.
-
Lentiviral-mediated delivery of mutant huntingtin in the striatum of rats induces a selective neuropathology modulated by polyglutamine repeat size, huntingtin expression levels, and protein length.J Neurosci. 2002 May 1;22(9):3473-83. doi: 10.1523/JNEUROSCI.22-09-03473.2002. J Neurosci. 2002. PMID: 11978824 Free PMC article.
-
Huntington's disease.Adv Exp Med Biol. 2010;685:45-63. Adv Exp Med Biol. 2010. PMID: 20687494 Review.
-
Transgenic animal models for study of the pathogenesis of Huntington's disease and therapy.Drug Des Devel Ther. 2015 Apr 15;9:2179-88. doi: 10.2147/DDDT.S58470. eCollection 2015. Drug Des Devel Ther. 2015. PMID: 25931812 Free PMC article. Review.
Cited by
-
Influence of species differences on the neuropathology of transgenic Huntington's disease animal models.J Genet Genomics. 2012 Jun 20;39(6):239-45. doi: 10.1016/j.jgg.2012.05.002. Epub 2012 May 14. J Genet Genomics. 2012. PMID: 22749010 Free PMC article. Review.
-
Small Ruminants and Its Use in Regenerative Medicine: Recent Works and Future Perspectives.Biology (Basel). 2021 Mar 22;10(3):249. doi: 10.3390/biology10030249. Biology (Basel). 2021. PMID: 33810087 Free PMC article. Review.
-
Sheep and Goat Genome Engineering: From Random Transgenesis to the CRISPR Era.Front Genet. 2019 Sep 3;10:750. doi: 10.3389/fgene.2019.00750. eCollection 2019. Front Genet. 2019. PMID: 31552084 Free PMC article. Review.
-
Artificial miRNAs Reduce Human Mutant Huntingtin Throughout the Striatum in a Transgenic Sheep Model of Huntington's Disease.Hum Gene Ther. 2018 Jun;29(6):663-673. doi: 10.1089/hum.2017.199. Epub 2018 Feb 23. Hum Gene Ther. 2018. PMID: 29207890 Free PMC article.
-
CRISPR/Cas9: a powerful genetic engineering tool for establishing large animal models of neurodegenerative diseases.Mol Neurodegener. 2015 Aug 4;10:35. doi: 10.1186/s13024-015-0031-x. Mol Neurodegener. 2015. PMID: 26238861 Free PMC article. Review.
References
-
- Huntington's Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. The Huntington's Disease Collaborative Research Group. Cell. 1993;72:971–983. - PubMed
-
- Mangiarini L., Sathasivam K., Seller M., Cozens B.A., Harper A., Hetherington C., Lawton M., Trottier Y., Lehrach H., Davies S.W., et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell. 1996;87:493–506. - PubMed
-
- Scheerlinck J.-P.Y., Snibson K.J., Bowles V.M., Sutton P. Biomedical applications of sheep models: from asthma to vaccines. Trends Biotechnol. 2008;26:259–266. - PubMed
-
- Adalsteinsson S., Basrur P.K. Inheritance of spina bifida in Icelandic lambs. J. Hered. 1984;75:378–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
