

Aminoglycoside activity observed on single pre-translocation ribosome complexes
- PMID: 20154669
- PMCID: PMC3566567
- DOI: 10.1038/nchembio0310-244c
Aminoglycoside activity observed on single pre-translocation ribosome complexes
Abstract
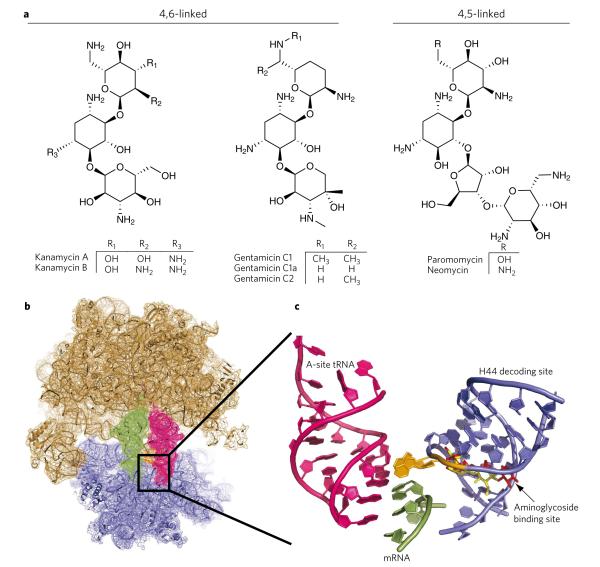
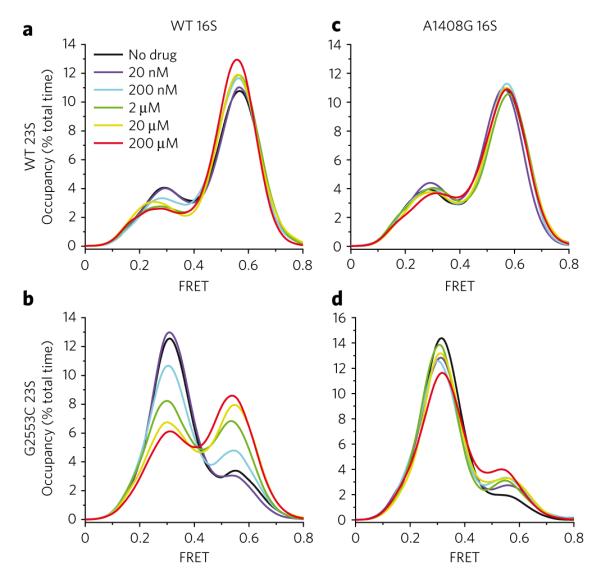
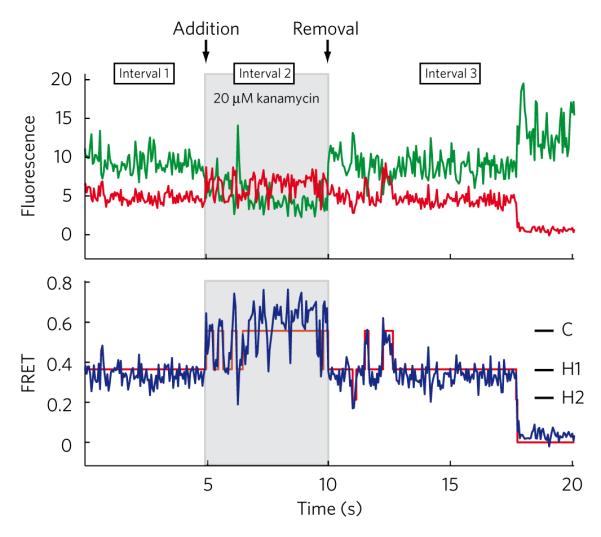
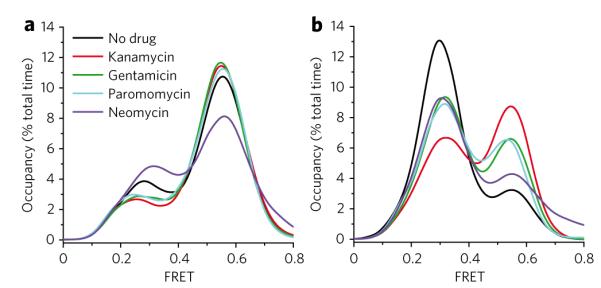
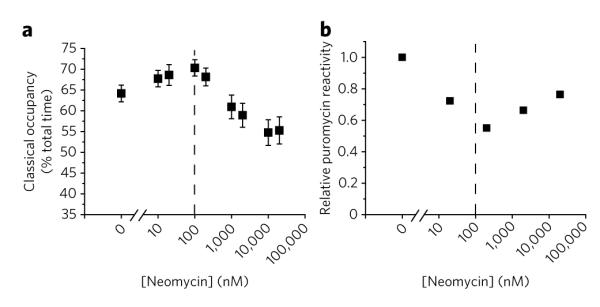
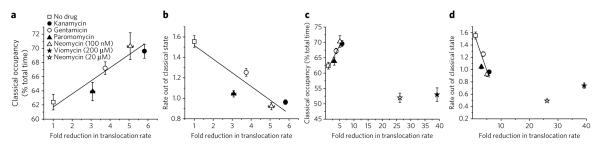
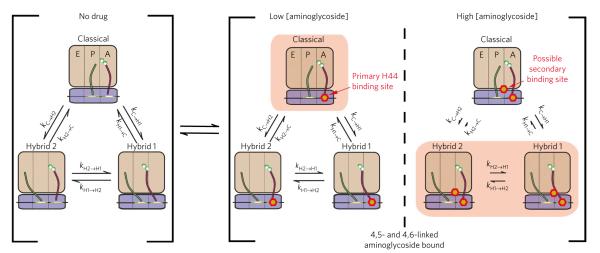
Aminoglycoside-class antibiotics bind directly to ribosomal RNA, imparting pleiotropic effects on ribosome function. Despite in-depth structural investigations of aminoglycoside–RNA oligonucleotide and aminoglycoside-ribosome interactions, mechanisms explaining the unique ribosome inhibition profiles of chemically similar aminoglycosides remain elusive. Here, using single-molecule fluorescence resonance energy transfer (smFRET) methods, we show that high-affinity aminoglycoside binding to the conserved decoding site region of the functional pre-translocation ribosome complex specifically remodels the nature of intrinsic dynamic processes within the particle. The extents of these effects, which are distinct for each member of the aminoglycoside class, strongly correlate with their inhibition of EF-G–catalyzed translocation. Neomycin, a 4,5-linked aminoglycoside, binds with lower affinity to one or more secondary binding sites, mediating distinct structural and dynamic perturbations that further enhance translocation inhibition. These new insights help explain why closely related aminoglycosides elicit pleiotropic translation activities and demonstrate the potential utility of smFRET as a tool for dissecting the mechanisms of antibiotic action.
Figures
References
-
- Rodnina MV, Gromadski KB, Kothe U, Wieden HJ. Recognition and selection of tRNA in translation. FEBS Lett. 2005;579:938–942. - PubMed
-
- Korostelev A, Trakhanov S, Laurberg M, Noller HF. Crystal structure of a 70S ribosome-tRNA complex reveals functional interactions and rearrangements. Cell. 2006;126:1065–1077. - PubMed
-
- Selmer M, et al. Structure of the 70S ribosome complexed with mRNA and tRNA. Science. 2006;313:1935–1942. - PubMed
-
- Yusupov MM, et al. Crystal structure of the ribosome at 5.5 angstrom resolution. Science. 2001;292:883–896. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
