

Abundance of type I toxin-antitoxin systems in bacteria: searches for new candidates and discovery of novel families
- PMID: 20156992
- PMCID: PMC2887945
- DOI: 10.1093/nar/gkq054
Abundance of type I toxin-antitoxin systems in bacteria: searches for new candidates and discovery of novel families
Abstract
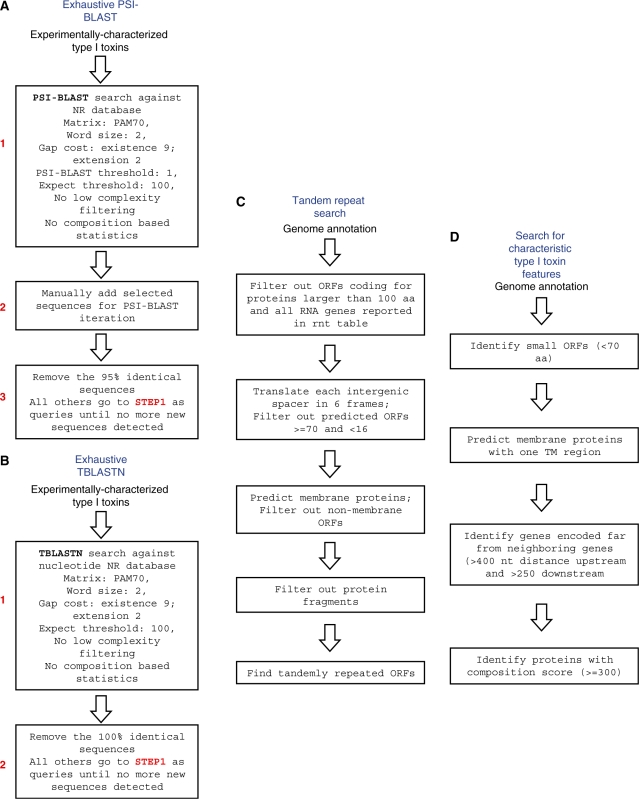
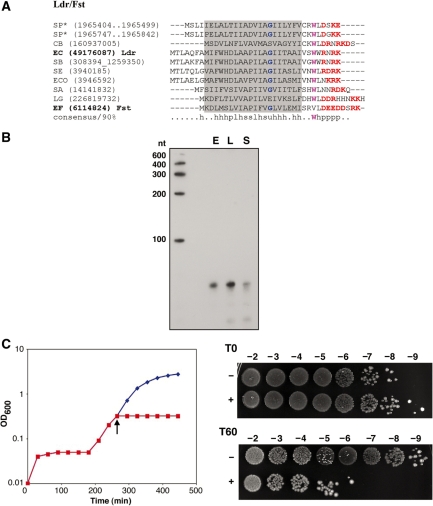
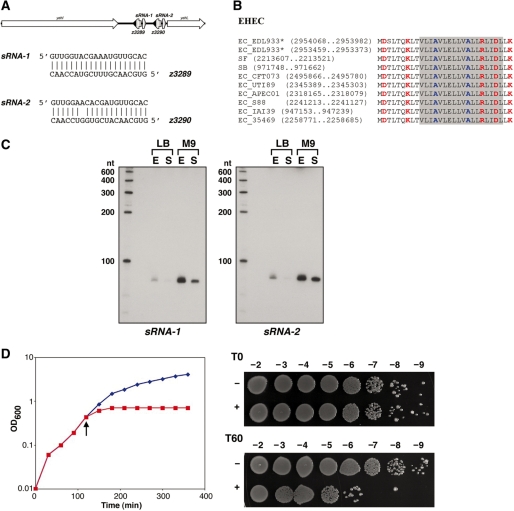
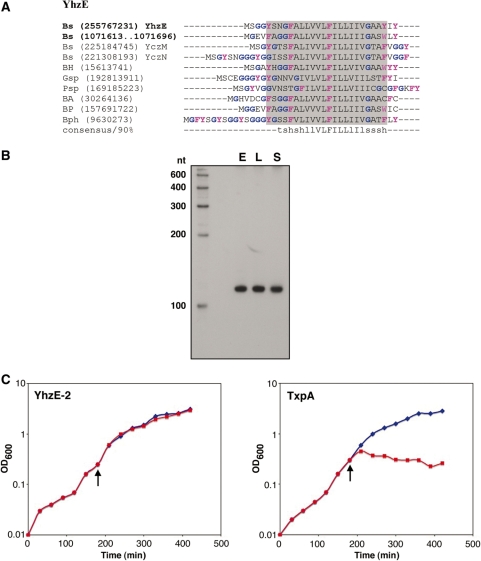
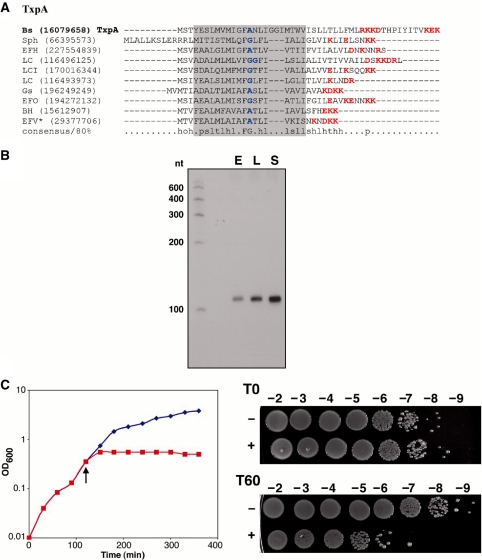
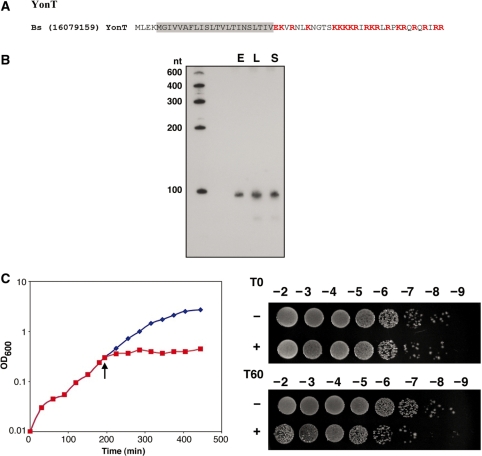
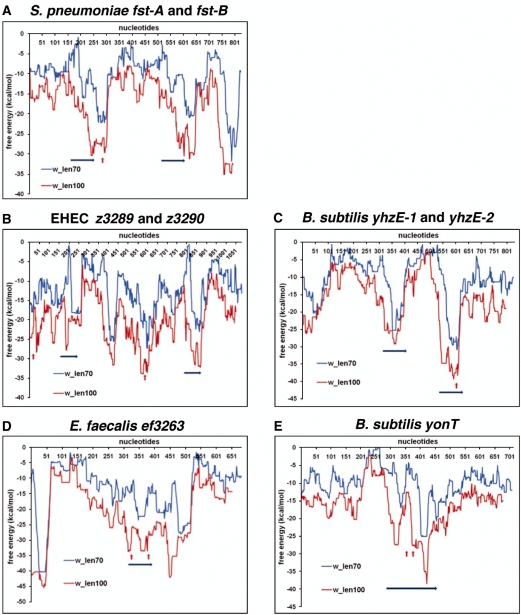
Small, hydrophobic proteins whose synthesis is repressed by small RNAs (sRNAs), denoted type I toxin-antitoxin modules, were first discovered on plasmids where they regulate plasmid stability, but were subsequently found on a few bacterial chromosomes. We used exhaustive PSI-BLAST and TBLASTN searches across 774 bacterial genomes to identify homologs of known type I toxins. These searches substantially expanded the collection of predicted type I toxins, revealed homology of the Ldr and Fst toxins, and suggested that type I toxin-antitoxin loci are not spread by horizontal gene transfer. To discover novel type I toxin-antitoxin systems, we developed a set of search parameters based on characteristics of known loci including the presence of tandem repeats and clusters of charged and bulky amino acids at the C-termini of short proteins containing predicted transmembrane regions. We detected sRNAs for three predicted toxins from enterohemorrhagic Escherichia coli and Bacillus subtilis, and showed that two of the respective proteins indeed are toxic when overexpressed. We also demonstrated that the local free-energy minima of RNA folding can be used to detect the positions of the sRNA genes. Our results suggest that type I toxin-antitoxin modules are much more widely distributed among bacteria than previously appreciated.
Figures
References
-
- Yamaguchi Y, Inouye M. mRNA interferases, sequence-specific endoribonucleases from the toxin-antitoxin systems. Prog. Mol. Biol. Transl. Sci. 2009;85:467–500. - PubMed
-
- Gerdes K, Christensen SK, Løbner-Olesen A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005;3:371–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
