

Structure prediction-driven genetics in Saccharomyces cerevisiae identifies an interface between the t-RPA proteins Stn1 and Ten1
- PMID: 20157006
- PMCID: PMC2870947
- DOI: 10.1534/genetics.109.111922
Structure prediction-driven genetics in Saccharomyces cerevisiae identifies an interface between the t-RPA proteins Stn1 and Ten1
Abstract
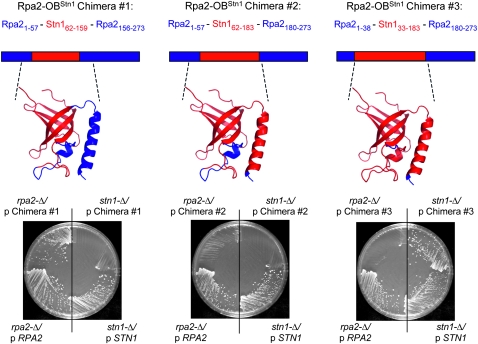
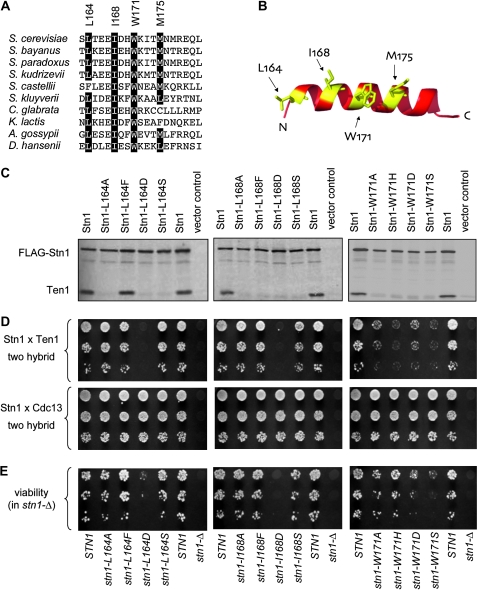
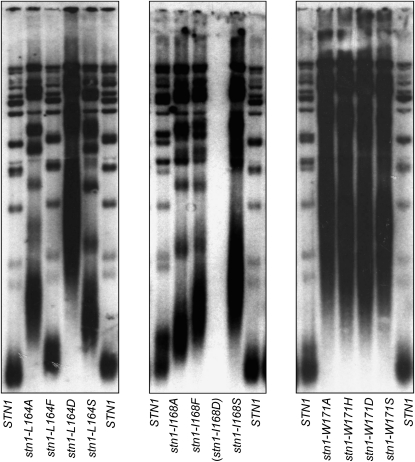
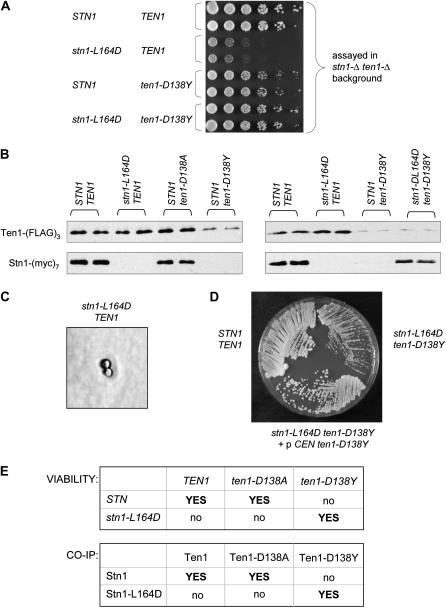
In Saccharomyces cerevisiae, Cdc13, Stn1, and Ten1 are essential for both chromosome capping and telomere length homeostasis. These three proteins have been proposed to perform their roles at chromosome termini as a telomere-dedicated t-RPA complex, on the basis of several parallels with the conventional RPA complex. In this study, we have used several approaches to test whether a predicted alpha-helix in the N-terminal domain of the S. cerevisiae Stn1 protein is required for formation of the proposed t-RPA complex, in a manner analogous to the comparable helix in Rpa2. Analysis of a panel of Rpa2-OB(Stn1) chimeras indicates that whether a chimeric protein contains the Rpa2 or Stn1 version of this alpha-helix dictates its ability to function in place of Rpa2 or Stn1, respectively. In addition, mutations introduced into a hydrophobic surface of the predicted Stn1 alpha-helix eliminated association with Ten1. Strikingly, allele-specific suppression of a stn1 mutation in this helix (stn1-L164D) by a ten1 mutation (ten1-D138Y) resulted in a restored Stn1-Ten1 interaction, supporting the identification of a Stn1-Ten1 interface. We conclude that Stn1 interacts with Ten1 through an alpha-helix, in a manner analogous to the interaction between the comparable subunits of the RPA complex.
Figures


Similar articles
-
RPA-like proteins mediate yeast telomere function.Nat Struct Mol Biol. 2007 Mar;14(3):208-14. doi: 10.1038/nsmb1205. Epub 2007 Feb 11. Nat Struct Mol Biol. 2007. PMID: 17293872
-
Distinct roles for yeast Stn1 in telomere capping and telomerase inhibition.EMBO J. 2008 Sep 3;27(17):2328-39. doi: 10.1038/emboj.2008.158. EMBO J. 2008. PMID: 19172739 Free PMC article.
-
Ten1 functions in telomere end protection and length regulation in association with Stn1 and Cdc13.EMBO J. 2001 Mar 1;20(5):1173-83. doi: 10.1093/emboj/20.5.1173. EMBO J. 2001. PMID: 11230140 Free PMC article.
-
Evolution of CST function in telomere maintenance.Cell Cycle. 2010 Aug 15;9(16):3157-65. doi: 10.4161/cc.9.16.12547. Epub 2010 Aug 26. Cell Cycle. 2010. PMID: 20697207 Free PMC article. Review.
-
RPA-like single-stranded DNA-binding protein complexes including CST serve as specialized processivity factors for polymerases.Curr Opin Struct Biol. 2023 Aug;81:102611. doi: 10.1016/j.sbi.2023.102611. Epub 2023 May 26. Curr Opin Struct Biol. 2023. PMID: 37245465 Free PMC article. Review.
Cited by
-
Arabidopsis ATM and ATR kinases prevent propagation of genome damage caused by telomere dysfunction.Plant Cell. 2011 Dec;23(12):4254-65. doi: 10.1105/tpc.111.092387. Epub 2011 Dec 9. Plant Cell. 2011. PMID: 22158468 Free PMC article.
-
Dimerization of Cdc13 is essential for dynamic DNA exchange on telomeric DNA.bioRxiv [Preprint]. 2025 Mar 26:2025.03.25.645294. doi: 10.1101/2025.03.25.645294. bioRxiv. 2025. Update in: J Biol Chem. 2025 Jul 17;301(8):110496. doi: 10.1016/j.jbc.2025.110496. PMID: 40196551 Free PMC article. Updated. Preprint.
-
Telomerase recruitment in Saccharomyces cerevisiae is not dependent on Tel1-mediated phosphorylation of Cdc13.Genetics. 2010 Dec;186(4):1147-59. doi: 10.1534/genetics.110.122044. Epub 2010 Sep 13. Genetics. 2010. PMID: 20837994 Free PMC article.
-
Analyses of Candida Cdc13 orthologues revealed a novel OB fold dimer arrangement, dimerization-assisted DNA binding, and substantial structural differences between Cdc13 and RPA70.Mol Cell Biol. 2012 Jan;32(1):186-98. doi: 10.1128/MCB.05875-11. Epub 2011 Oct 24. Mol Cell Biol. 2012. PMID: 22025677 Free PMC article.
-
Tying up the Ends: Plasticity in the Recognition of Single-Stranded DNA at Telomeres.Biochemistry. 2016 Sep 27;55(38):5326-40. doi: 10.1021/acs.biochem.6b00496. Epub 2016 Sep 15. Biochemistry. 2016. PMID: 27575340 Free PMC article.
References
-
- Bianchi, A., S. Negrini and D. Shore, 2004. Delivery of yeast telomerase to a DNA break depends on the recruitment functions of Cdc13 and Est1. Mol. Cell 16 139–146. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
