

Molecular basis for an attenuated mitochondrial adaptive plasticity in aged skeletal muscle
- PMID: 20157569
- PMCID: PMC2815739
- DOI: 10.18632/aging.100083
Molecular basis for an attenuated mitochondrial adaptive plasticity in aged skeletal muscle
Abstract
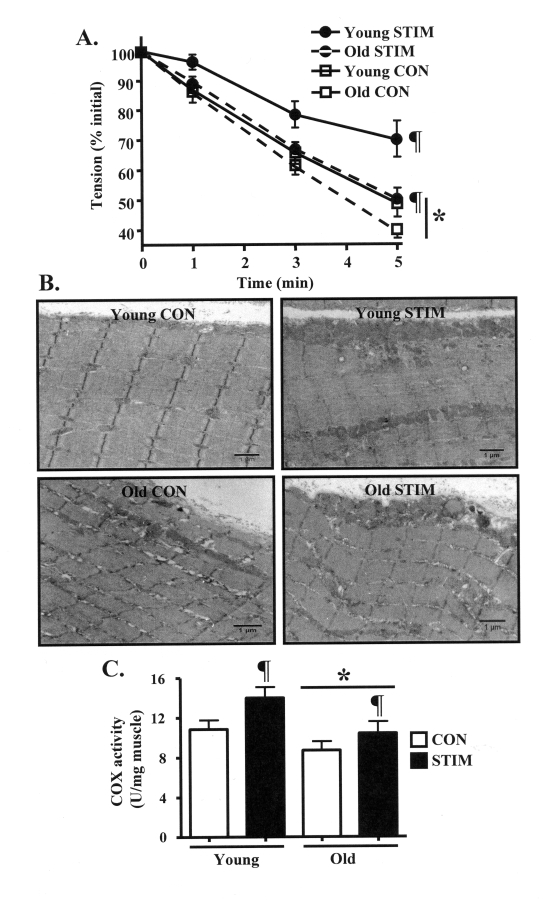
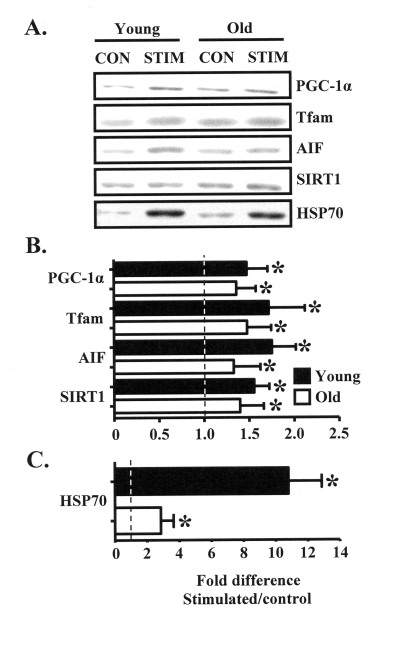
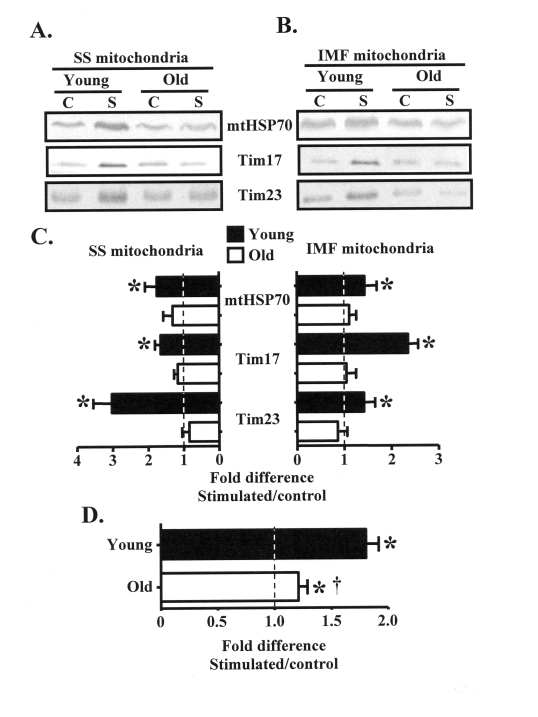
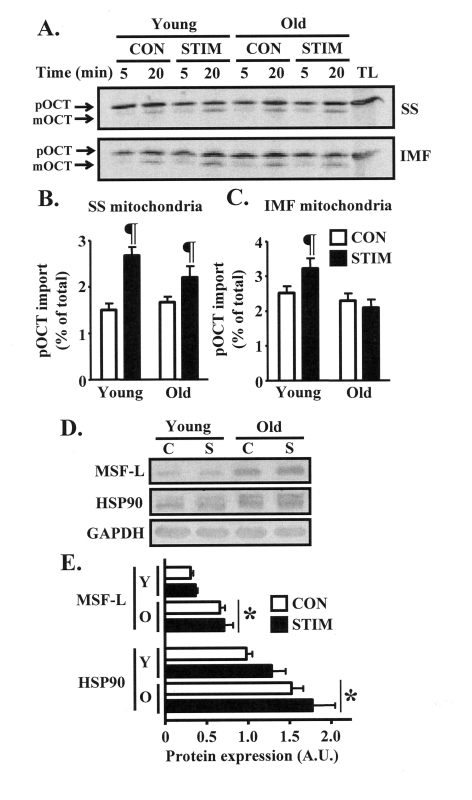
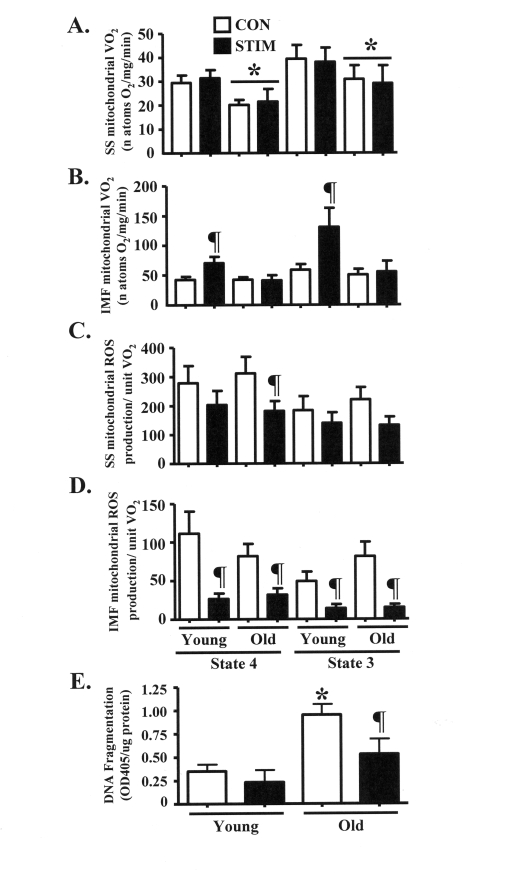
Our intent was to investigate the mechanisms driving the adaptive potential of subsarcolemmal (SS) and intermyofibrillar (IMF) mitochondria in young (6 mo) and senescent (36 mo) animals in response to a potent stimulus for organelle biogenesis. We employed chronic electrical stimulation (10 Hz, 3 h/day, 7 days) to induce contractile activity of skeletal muscle in 6 and 36 mo F344XBN rats. Subsequent to chronic activity, acute stimulation (1 Hz, 5 min) in situ revealed greater fatigue resistance in both age groups. However, the improvement in endurance was significantly greater in the young, compared to the old animals. Chronic muscle use also augmented SS and IMF mitochondrial volume to a greater extent in young muscle. The molecular basis for the diminished organelle expansion in aged muscle was due, in part, to the collective attenuation of the chronic stimulation-evoked increase in regulatory proteins involved in mediating mitochondrial protein import and biogenesis. Furthermore, adaptations in mitochondrial function were also blunted in old animals. However, chronic contractile activity evoked greater reductions in mitochondrially-mediated proapoptotic signaling in aged muscle. Thus, mitochondrial plasticity is retained in aged animals, however the magnitude of the changes are less compared to young animals due to attenuated molecular processes regulating organelle biogenesis.
Keywords: apoptosis; chronic contractile activity; intermyofibrillar mitochondria; performance; protein import; subsarcolemmal mitochondria.
Conflict of interest statement
The authors declare no conflict of interests.
Figures
Similar articles
-
Effect of chronic contractile activity on SS and IMF mitochondrial apoptotic susceptibility in skeletal muscle.Am J Physiol Endocrinol Metab. 2007 Mar;292(3):E748-55. doi: 10.1152/ajpendo.00311.2006. Epub 2006 Nov 14. Am J Physiol Endocrinol Metab. 2007. PMID: 17106065
-
Mitochondrial function and apoptotic susceptibility in aging skeletal muscle.Aging Cell. 2008 Jan;7(1):2-12. doi: 10.1111/j.1474-9726.2007.00347.x. Epub 2007 Nov 19. Aging Cell. 2008. PMID: 18028258
-
Effect of contractile activity on protein turnover in skeletal muscle mitochondrial subfractions.J Appl Physiol (1985). 2000 May;88(5):1601-6. doi: 10.1152/jappl.2000.88.5.1601. J Appl Physiol (1985). 2000. PMID: 10797119
-
Mitochondrial assembly: protein import.Proc Nutr Soc. 2004 May;63(2):293-300. doi: 10.1079/PNS2004342. Proc Nutr Soc. 2004. PMID: 15294046 Review.
-
Plasticity of skeletal muscle mitochondria in response to contractile activity.Exp Physiol. 2003 Jan;88(1):99-107. doi: 10.1113/eph8802505. Exp Physiol. 2003. PMID: 12525859 Review.
Cited by
-
Mitochondrial Quantity and Quality in Age-Related Sarcopenia.Int J Mol Sci. 2024 Feb 8;25(4):2052. doi: 10.3390/ijms25042052. Int J Mol Sci. 2024. PMID: 38396729 Free PMC article. Review.
-
Regulation of satellite cell function in sarcopenia.Front Aging Neurosci. 2014 Sep 22;6:246. doi: 10.3389/fnagi.2014.00246. eCollection 2014. Front Aging Neurosci. 2014. PMID: 25295003 Free PMC article. Review.
-
Mitochondrial Biogenesis in Skeletal Muscle.Adv Exp Med Biol. 2025;1478:19-50. doi: 10.1007/978-3-031-88361-3_2. Adv Exp Med Biol. 2025. PMID: 40879934 Review.
-
MAP kinase phosphatase-1--a new player at the nexus between sarcopenia and metabolic disease.Aging (Albany NY). 2010 Apr 6;2(3):170-6. doi: 10.18632/aging.100135. Aging (Albany NY). 2010. PMID: 20375469 Free PMC article. Review.
-
Contractile activity attenuates autophagy suppression and reverses mitochondrial defects in skeletal muscle cells.Autophagy. 2018;14(11):1886-1897. doi: 10.1080/15548627.2018.1491488. Epub 2018 Aug 4. Autophagy. 2018. PMID: 30078345 Free PMC article.
References
-
- Hood DA. Invited Review: contractile activity-induced mitochondrial biogenesis in skeletal muscle. J Appl Physiol. 2001;90:1137–1157. - PubMed
-
- Akimoto T, Pohnert SC, Li P, Zhang M, Gumbs C, Rosenberg PB, Williams RS, Yan Z. Exercise stimulates Pgc-1alpha transcription in skeletal muscle through activation of the p38 MAPK pathway. J Biol Chem. 2005;280:19587–19593. - PubMed
-
- Wright DC, Geiger PC, Han DH, Jones TE, Holloszy JO. Calcium induces increases in peroxisome proliferator-activated receptor gamma coactivator-1alpha and mitochondrial biogenesis by a pathway leading to p38 mitogen-activated protein kinase activation. J Biol Chem. 2007;282:18793–18799. - PubMed
-
- Hoppeler H. Exercise-induced ultrastructural changes in skeletal muscle. Int J Sports Med. 1986;7:187–204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical