

Sonic hedgehog signaling confers ventral telencephalic progenitors with distinct cortical interneuron fates
- PMID: 20159447
- PMCID: PMC2868511
- DOI: 10.1016/j.neuron.2010.01.004
Sonic hedgehog signaling confers ventral telencephalic progenitors with distinct cortical interneuron fates
Abstract
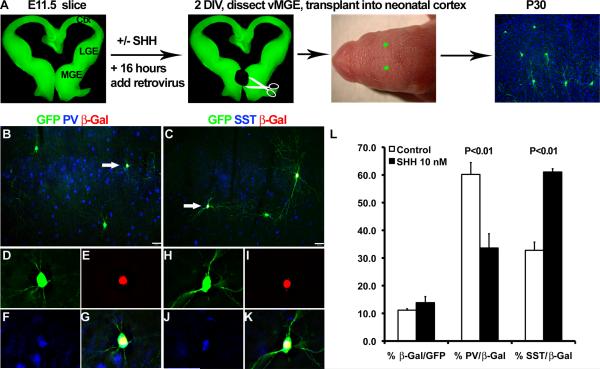
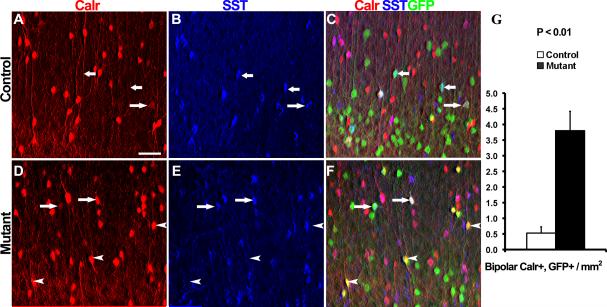
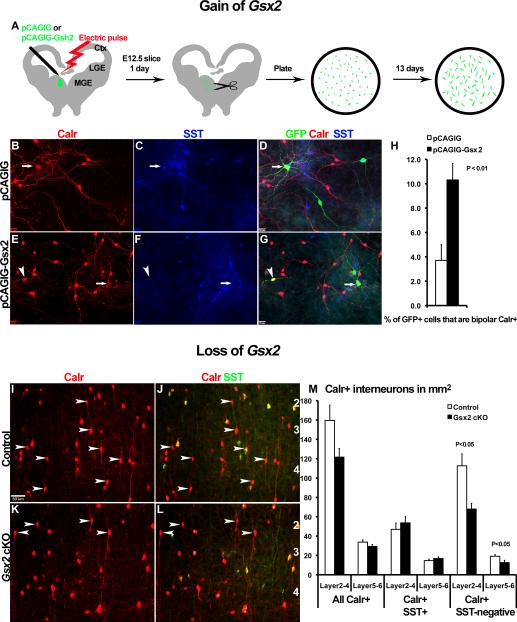
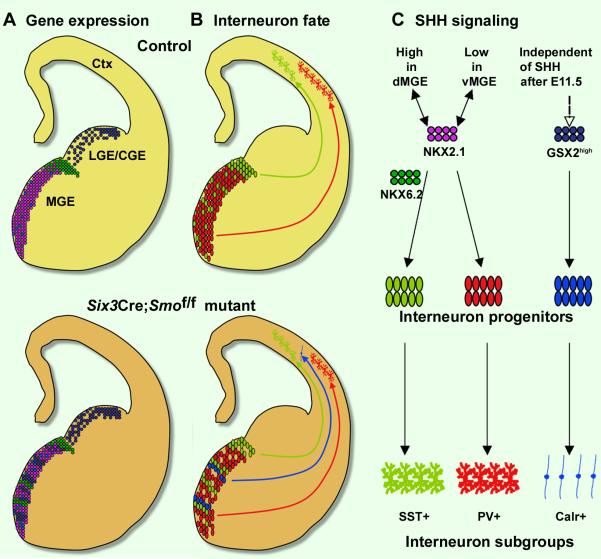
Interneurons in the cerebral cortex regulate cortical functions through the actions of distinct subgroups that express parvalbumin, somatostatin, or calretinin. The genesis of the first two subgroups requires the expression of NKX2.1, which is maintained by SHH signaling during neurogenesis. In this paper, we report that mosaic elimination in the medial ganglionic eminence (MGE) of Smo, a key effector of SHH signaling, reveals that MGE progenitors retain a remarkable degree of plasticity during the neurogenic period. SHH signaling prevents the upregulation of GSX2 and conversion of some MGE progenitors to a caudal ganglionic eminence-like, bipolar calretinin-expressing cell fate that is promoted by GSX2. In addition, a higher level of SHH signaling promotes the generation of the somatostatin-expressing interneuron at the expense of parvalbumin-expressing subgroup. These results indicate that cortical interneuron diversity, a major determinant of cortical function, is critically influenced by differential levels of SHH signaling within the ventral telencephalon.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
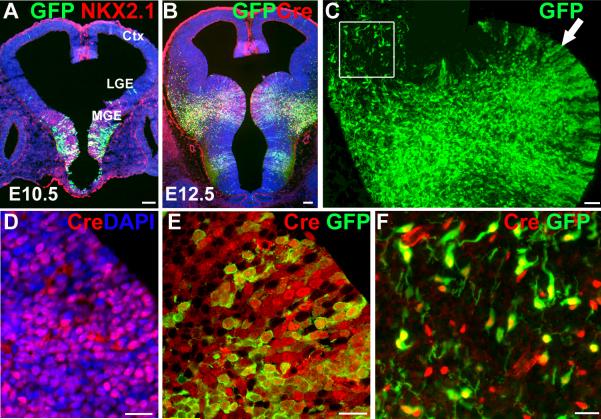
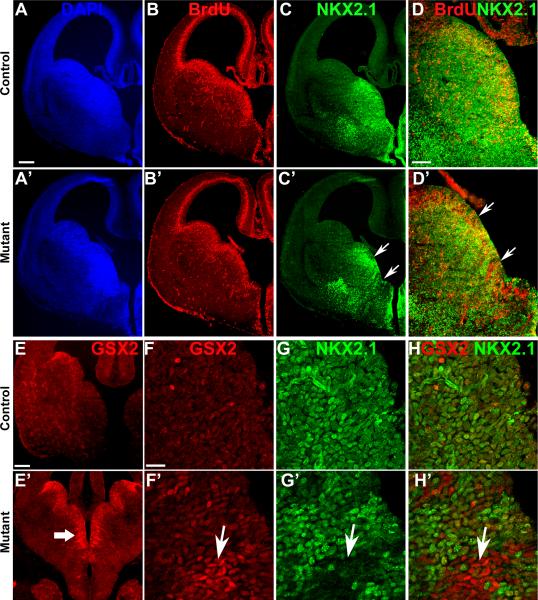
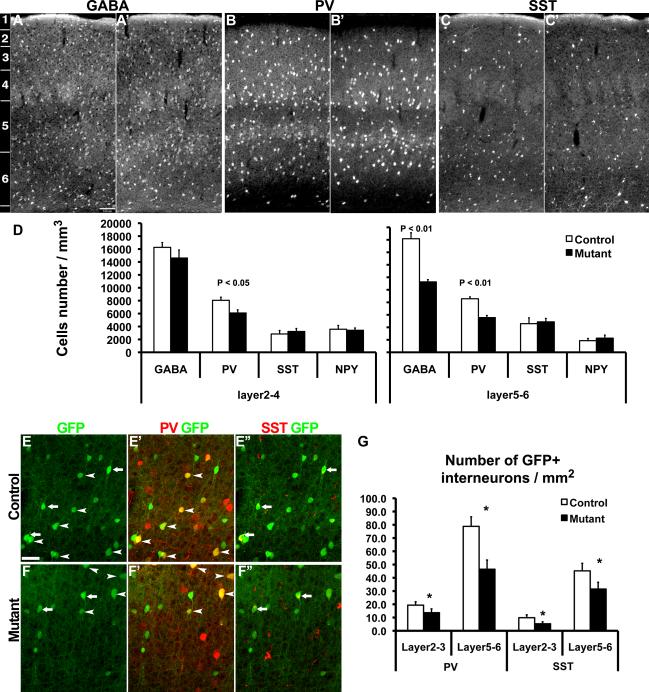
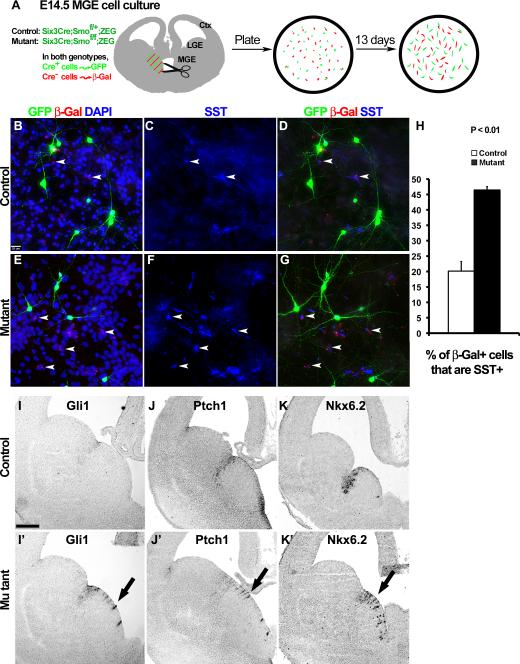
References
-
- Ahn S, Joyner AL. Dynamic changes in the response of cells to positive hedgehog signaling during mouse limb patterning. Cell. 2004;118:505–516. - PubMed
-
- Anderson SA, Marin O, Horn C, Jennings K, Rubenstein JL. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development. 2001;128:353–363. - PubMed
-
- Butt SJ, Fuccillo M, Nery S, Noctor S, Kriegstein A, Corbin JG, Fishell G. The temporal and spatial origins of cortical interneurons predict their physiological subtype. Neuron. 2005;48:591–604. - PubMed
-
- Cavanagh ME, Parnavelas JG. Development of vasoactive-intestinal-polypeptide-immunoreactive neurons in the rat occipital cortex: a combined immunohistochemical-autoradiographic study. J Comp Neurol. 1989;284:637–645. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
