

Regulation of synaptic structure and function by FMRP-associated microRNAs miR-125b and miR-132
- PMID: 20159450
- PMCID: PMC5018398
- DOI: 10.1016/j.neuron.2010.01.005
Regulation of synaptic structure and function by FMRP-associated microRNAs miR-125b and miR-132
Erratum in
- Neuron. 2010 Oct 6;68(1):161
Abstract
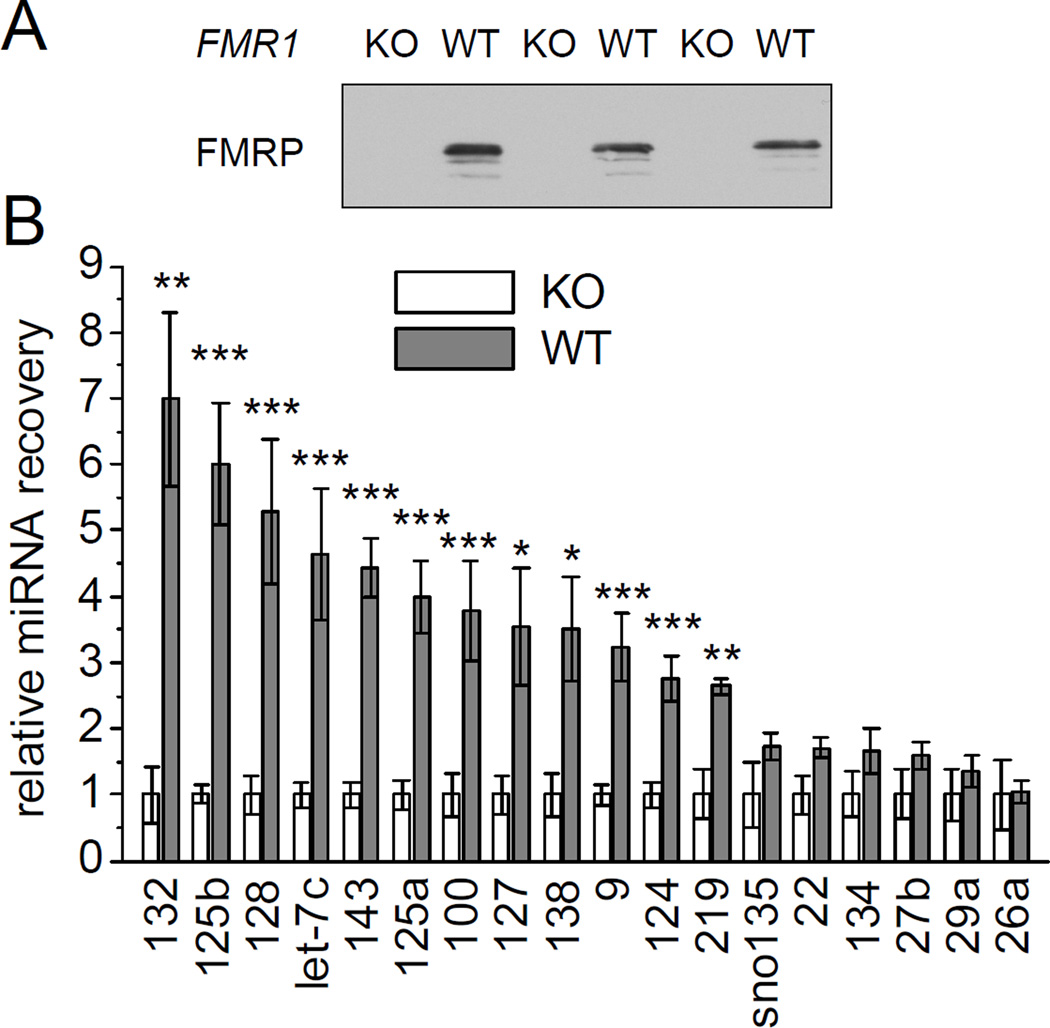
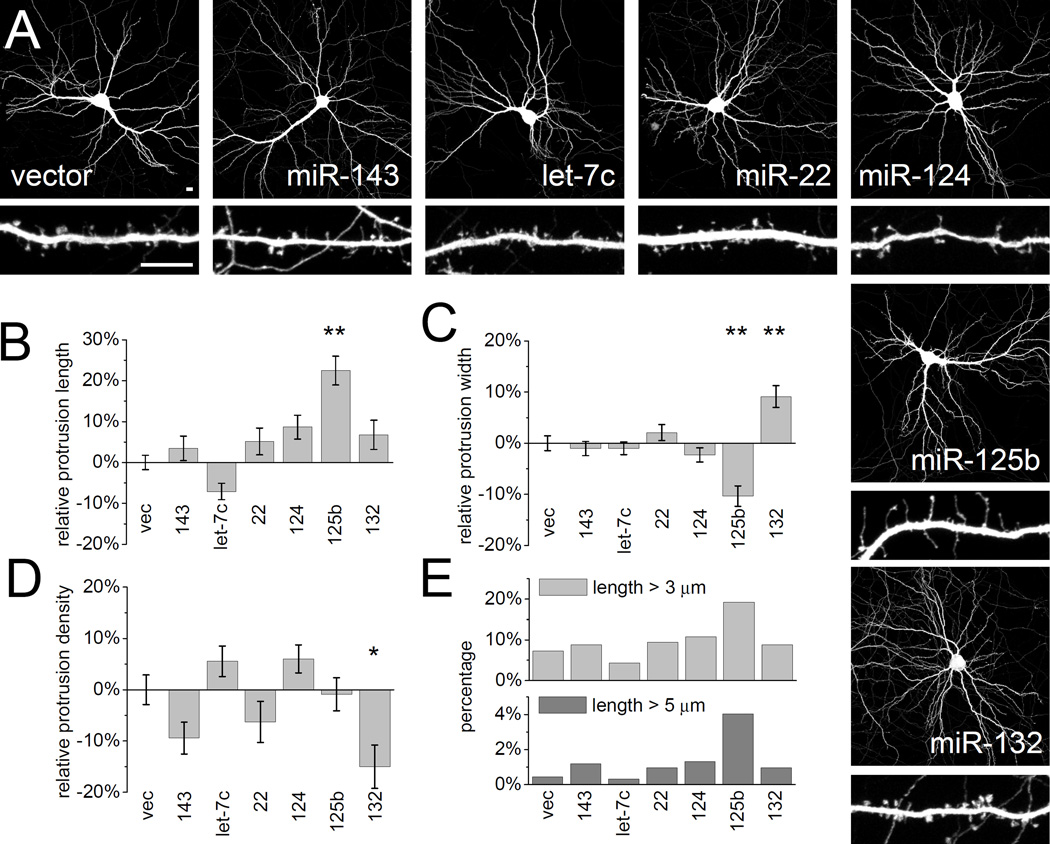
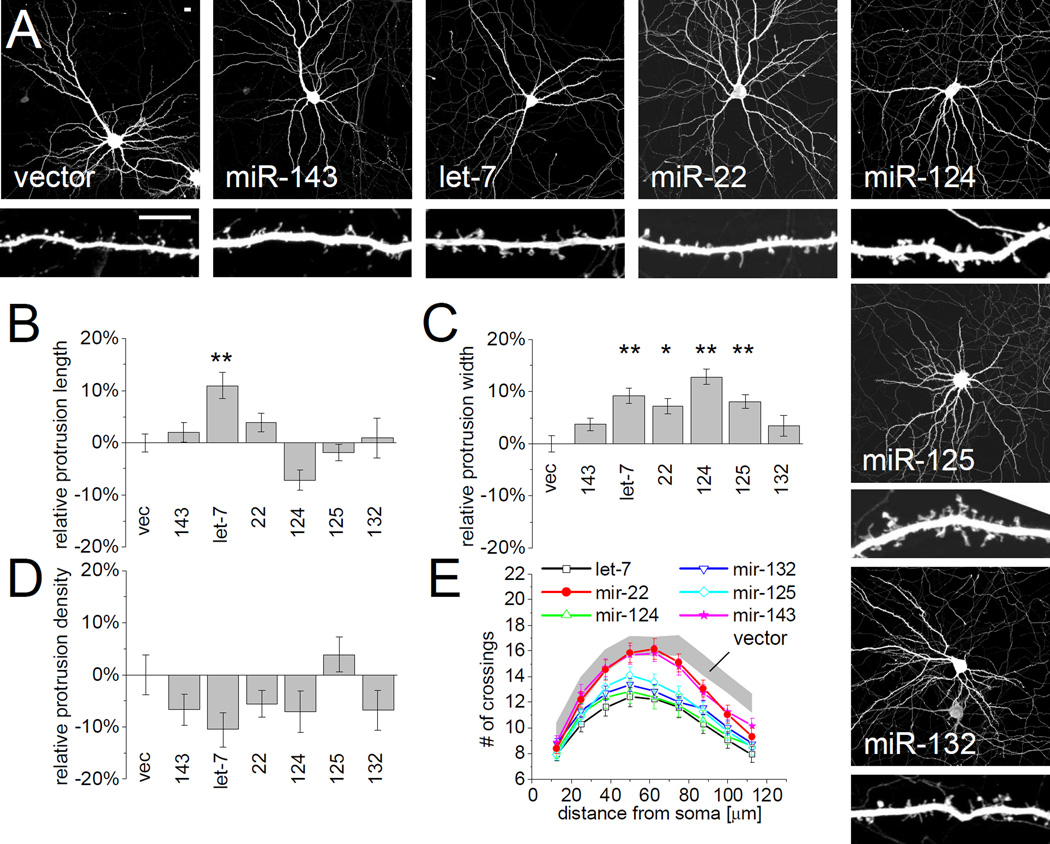
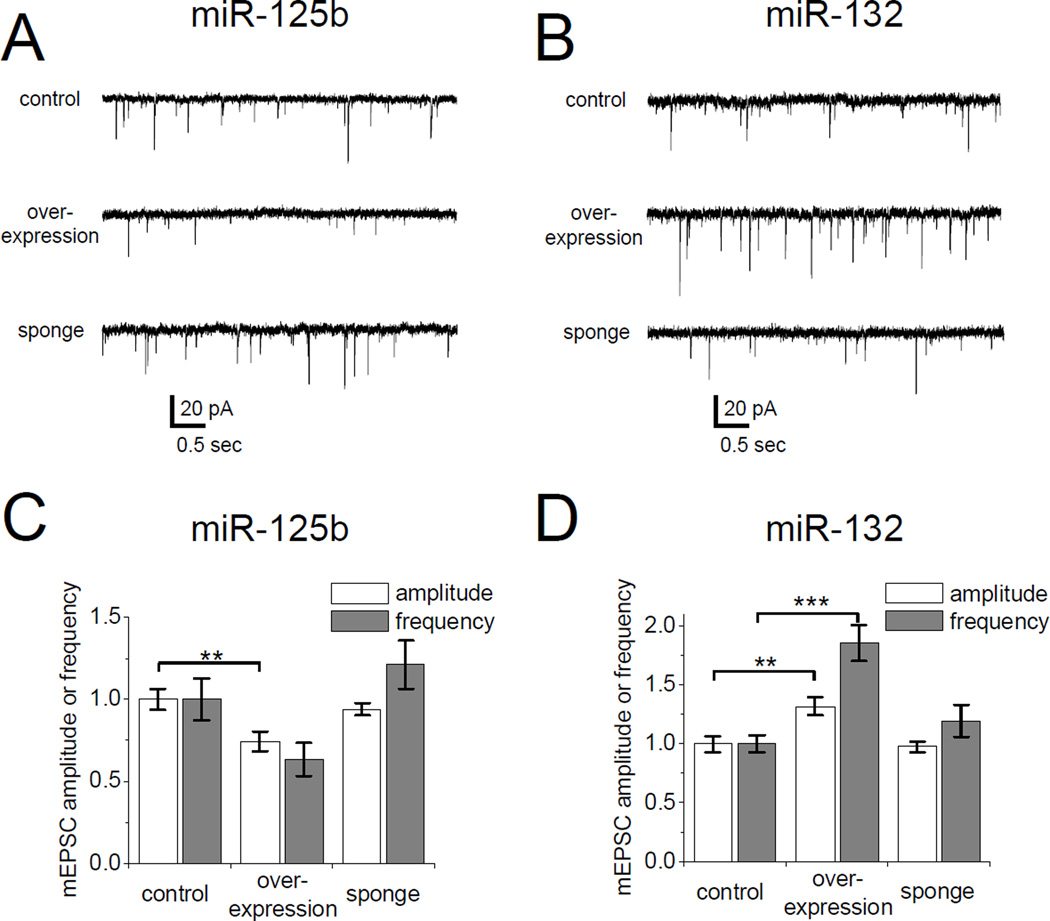
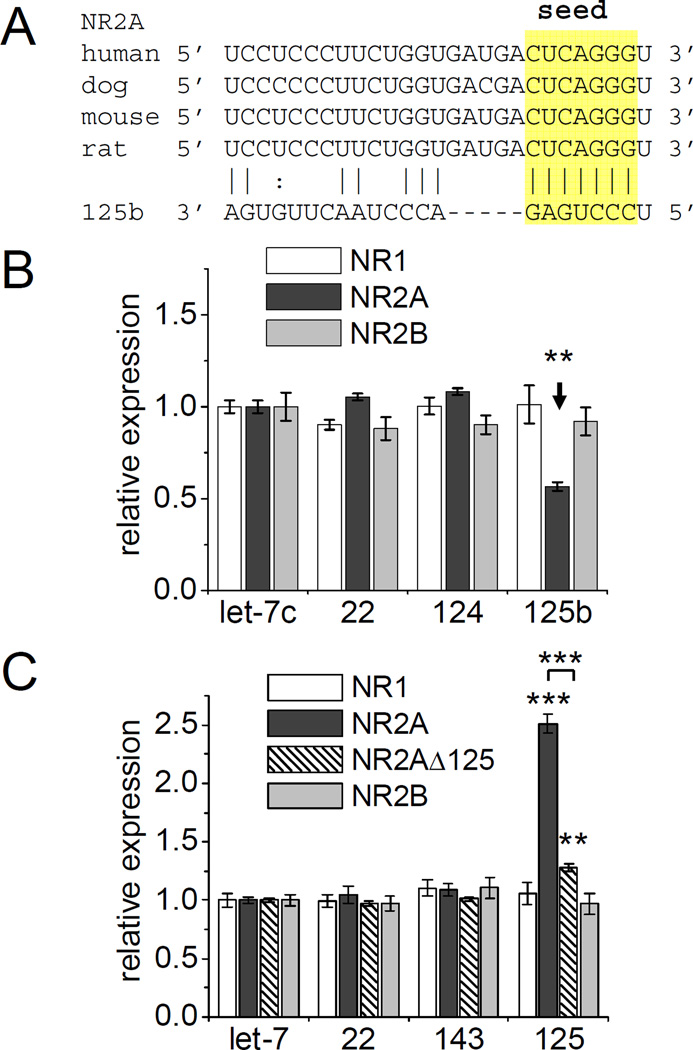
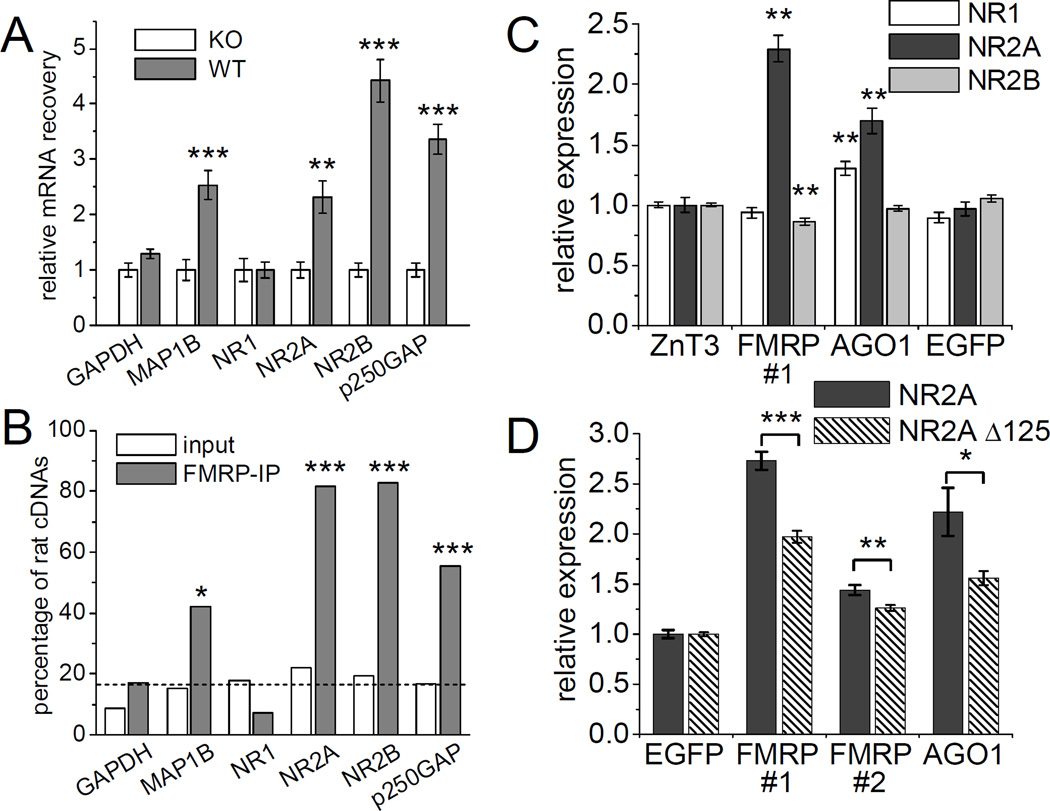
MicroRNAs (miRNAs) are noncoding RNAs that suppress translation of specific mRNAs. The miRNA machinery interacts with fragile X mental retardation protein (FMRP), which functions as translational repressor. We show that miR-125b and miR-132, as well as several other miRNAs, are associated with FMRP in mouse brain. miR-125b and miR-132 had largely opposing effects on dendritic spine morphology and synaptic physiology in hippocampal neurons. FMRP knockdown ameliorates the effect of miRNA overexpression on spine morphology. We identified NMDA receptor subunit NR2A as a target of miR-125b and show that NR2A mRNA is specifically associated with FMRP in brain. In hippocampal neurons, NR2A expression is negatively regulated through its 3' UTR by FMRP, miR-125b, and Argonaute 1. Regulation of NR2A 3'UTR by FMRP depends in part on miR-125b. Because NMDA receptor subunit composition profoundly affects synaptic plasticity, these observations have implications for the pathophysiology of fragile X syndrome, in which plasticity is altered.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
