

Structural basis of selective ubiquitination of TRF1 by SCFFbx4
- PMID: 20159592
- PMCID: PMC6097615
- DOI: 10.1016/j.devcel.2010.01.007
Structural basis of selective ubiquitination of TRF1 by SCFFbx4
Abstract
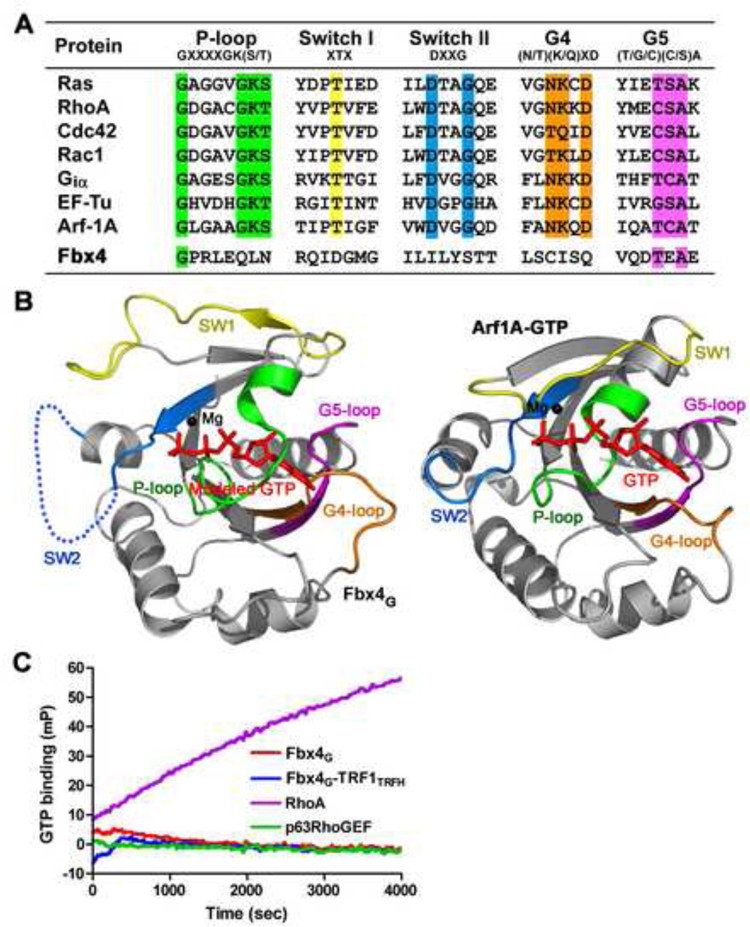
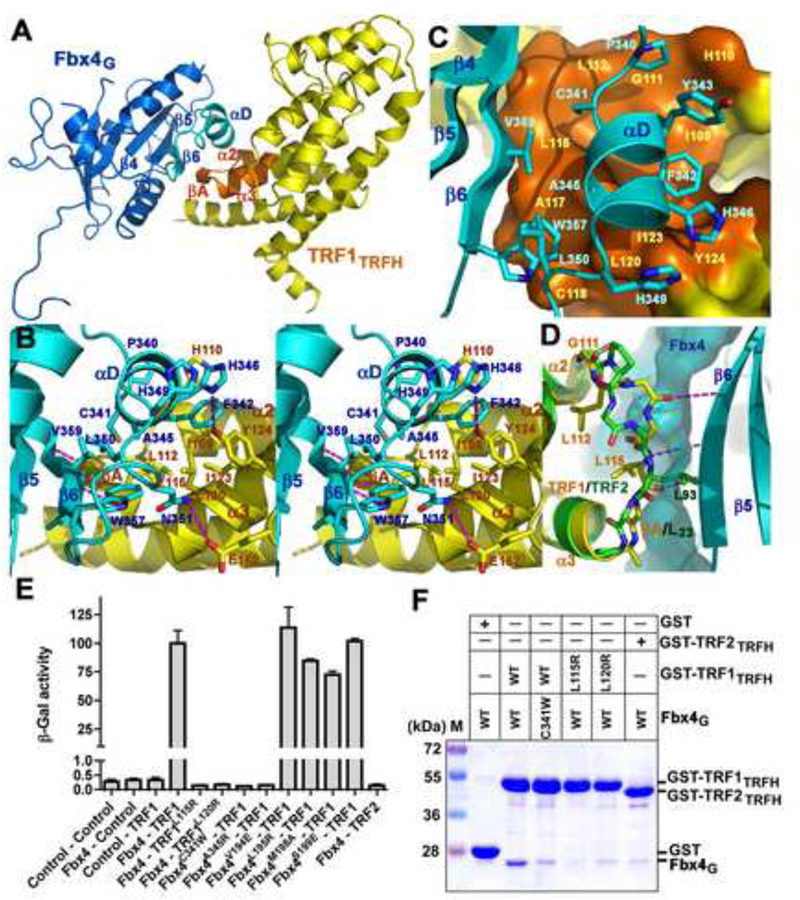
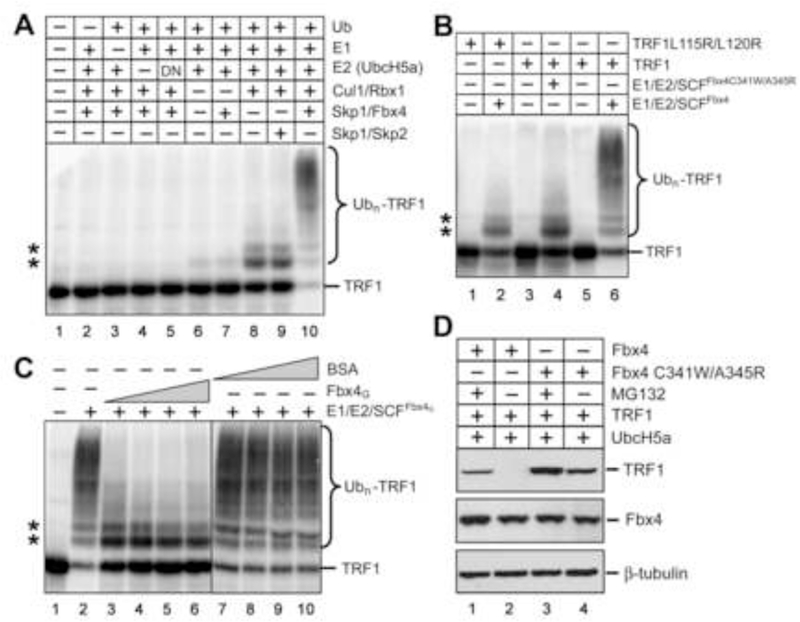
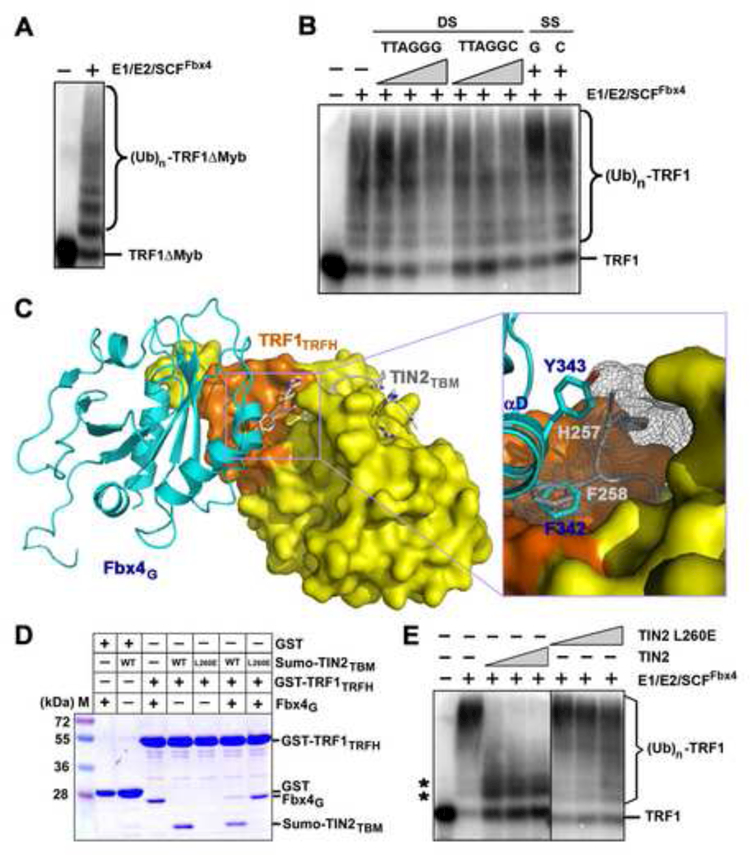
TRF1 is a critical regulator of telomere length. As such, TRF1 levels are regulated by ubiquitin-dependent proteolysis via an SCF E3 ligase where Fbx4 contributes to substrate specification. Here, we report the crystal structure of the Fbx4-TRF1 complex at 2.4 A resolution. Fbx4 contains an unusual substrate-binding domain that adopts a small GTPase fold. Strikingly, this atypical GTPase domain of Fbx4 binds to a globular domain of TRF1 through an intermolecular beta sheet, instead of recognizing short peptides/degrons as often seen in other F-box protein-substrate complexes. Importantly, mutations in this interface abrogate Fbx4-dependent TRF1 binding and ubiquitination. Furthermore, the data demonstrate that recognition of TRF1 by SCF(Fbx4) is regulated by another telomere protein, TIN2. Our results reveal an atypical small GTPase domain within Fbx4 as a substrate-binding motif for SCF(Fbx4) and uncover a mechanism for selective ubiquitination and degradation of TRF1 in telomere homeostasis control.
Copyright (c) 2010 Elsevier Inc. All rights reserved.
Figures

Comment in
-
Telomeric TuRF1 wars.Dev Cell. 2010 Feb 16;18(2):167-8. doi: 10.1016/j.devcel.2010.02.002. Dev Cell. 2010. PMID: 20159586
References
-
- Aspenstrom P, Ruusala A, and Pacholsky D (2007). Taking Rho GTPases to the next level: the cellular functions of atypical Rho GTPases. Experimental Cell Research 313, 3673–3679. - PubMed
-
- Blackburn EH (2001). Switching and signaling at the telomere. Cell 106, 661–673. - PubMed
-
- Broccoli D, Smogorzewska A, Chong L, and de Lange T (1997). Human telomeres contain two distinct Myb-related proteins, TRF1 and TRF2. Nature Genetics 17, 231–235. - PubMed
-
- Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, et al. (1998). Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr 54 (Pt 5), 905–921. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
