

Kinetochores generate microtubules with distal plus ends: their roles and limited lifetime in mitosis
- PMID: 20159595
- PMCID: PMC2828607
- DOI: 10.1016/j.devcel.2009.12.018
Kinetochores generate microtubules with distal plus ends: their roles and limited lifetime in mitosis
Abstract
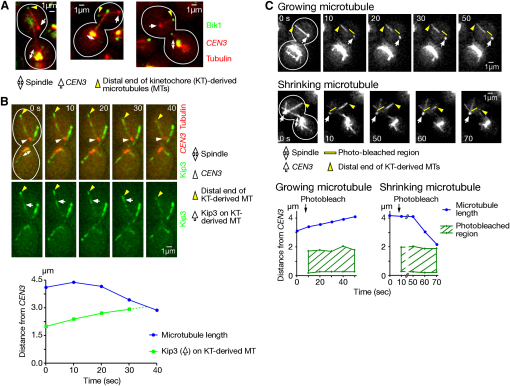
In early mitosis, microtubules can be generated at kinetochores as well as at spindle poles. However, the role and regulation of kinetochore-derived microtubules have been unclear. In general, metaphase spindle microtubules are oriented such that their plus ends bind to kinetochores. However, we now have evidence that, during early mitosis in budding yeast, microtubules are generated at kinetochores with distal plus ends. These kinetochore-derived microtubules interact along their length with microtubules that extend from a spindle pole, facilitating kinetochore loading onto the lateral surface of spindle pole microtubules. Once kinetochores are loaded, microtubules are no longer generated at kinetochores, and those that remain disappear rapidly and do not contribute to the metaphase spindle. Stu2 (the ortholog of vertebrate XMAP215/ch-TOG) localizes to kinetochores and plays a central role in regulating kinetochore-derived microtubules. Our work provides insight into microtubule generation at kinetochores and the mechanisms that facilitate initial kinetochore interaction with spindle pole microtubules.
Copyright (c) 2010 Elsevier Inc. All rights reserved.
Figures







Similar articles
-
Stu2 acts as a microtubule destabilizer in metaphase budding yeast spindles.Mol Biol Cell. 2018 Feb 1;29(3):247-255. doi: 10.1091/mbc.E17-08-0494. Epub 2017 Nov 29. Mol Biol Cell. 2018. PMID: 29187578 Free PMC article.
-
Molecular mechanisms of kinetochore capture by spindle microtubules.Nature. 2005 Apr 21;434(7036):987-94. doi: 10.1038/nature03483. Nature. 2005. PMID: 15846338
-
Kinetochore-dependent microtubule rescue ensures their efficient and sustained interactions in early mitosis.Dev Cell. 2011 Nov 15;21(5):920-33. doi: 10.1016/j.devcel.2011.09.006. Dev Cell. 2011. PMID: 22075150 Free PMC article.
-
Factors that Control Mitotic Spindle Dynamics.Adv Exp Med Biol. 2017;925:89-101. doi: 10.1007/5584_2016_74. Adv Exp Med Biol. 2017. PMID: 27722958 Review.
-
The composition, functions, and regulation of the budding yeast kinetochore.Genetics. 2013 Aug;194(4):817-46. doi: 10.1534/genetics.112.145276. Genetics. 2013. PMID: 23908374 Free PMC article. Review.
Cited by
-
Regulatory mechanisms of kinetochore-microtubule interaction in mitosis.Cell Mol Life Sci. 2013 Feb;70(4):559-79. doi: 10.1007/s00018-012-1057-7. Epub 2012 Jul 4. Cell Mol Life Sci. 2013. PMID: 22752158 Free PMC article. Review.
-
A stochastic model of kinetochore-microtubule attachment accurately describes fission yeast chromosome segregation.J Cell Biol. 2012 Mar 19;196(6):757-74. doi: 10.1083/jcb.201107124. Epub 2012 Mar 12. J Cell Biol. 2012. PMID: 22412019 Free PMC article.
-
Fibrous corona is reduced in cancer cell lines that attenuate microtubule nucleation from kinetochores.Cancer Sci. 2025 Feb;116(2):420-431. doi: 10.1111/cas.16406. Epub 2024 Nov 27. Cancer Sci. 2025. PMID: 39604214 Free PMC article.
-
Regulation of microtubule dynamics by TOG-domain proteins XMAP215/Dis1 and CLASP.Trends Cell Biol. 2011 Oct;21(10):604-14. doi: 10.1016/j.tcb.2011.06.007. Epub 2011 Jul 23. Trends Cell Biol. 2011. PMID: 21782439 Free PMC article. Review.
-
Biophysics of mitosis.Q Rev Biophys. 2012 May;45(2):147-207. doi: 10.1017/S0033583512000017. Epub 2012 Feb 10. Q Rev Biophys. 2012. PMID: 22321376 Free PMC article. Review.
References
-
- Akhmanova A., Steinmetz M.O. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat. Rev. Mol. Cell Biol. 2008;9:309–322. - PubMed
-
- Athale C.A., Dinarina A., Mora-Coral M., Pugieux C., Nedelec F., Karsenti E. Regulation of microtubule dynamics by reaction cascades around chromosomes. Science. 2008;322:1243–1247. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
