

Contribution of carbon fixed by Rubisco and PEPC to phloem export in the Crassulacean acid metabolism plant Kalanchoe daigremontiana
- PMID: 20159885
- PMCID: PMC2837257
- DOI: 10.1093/jxb/erq006
Contribution of carbon fixed by Rubisco and PEPC to phloem export in the Crassulacean acid metabolism plant Kalanchoe daigremontiana
Abstract
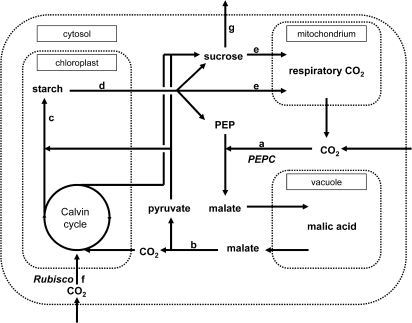
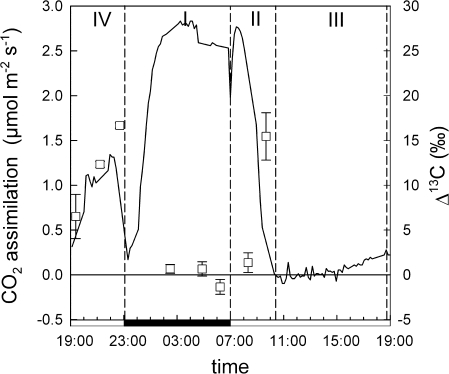
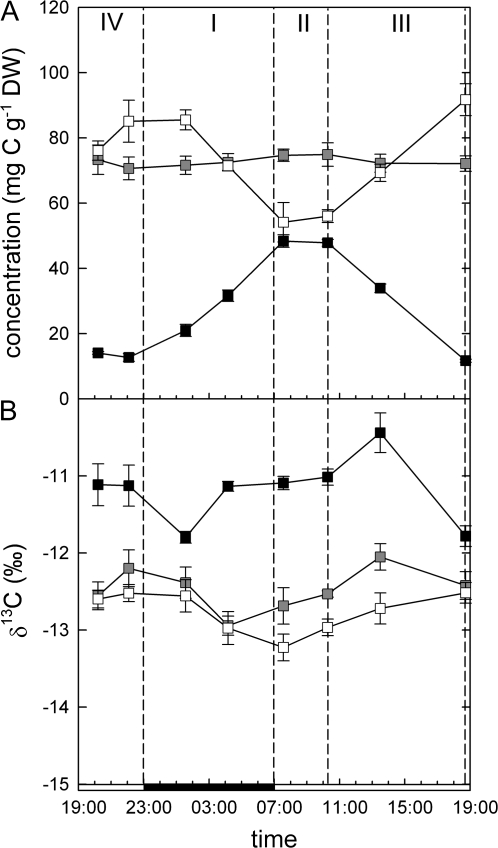
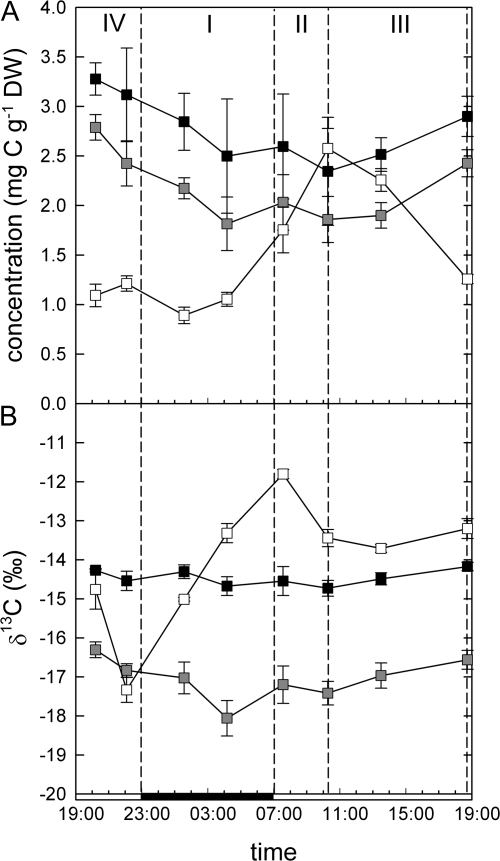
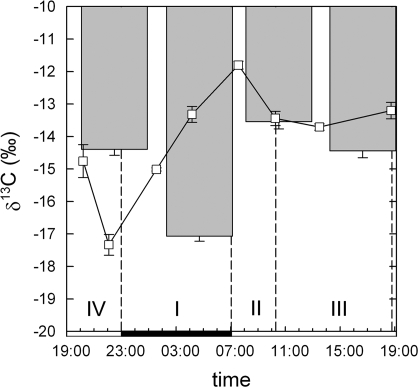
Crassulacean acid metabolism (CAM) plants exhibit a complex interplay between CO(2) fixation by phosphoenolpyruvate carboxylase (PEPC) and ribulose-1,5-bisphosphate carboxylase oxygenase (Rubisco), and carbon demand for CAM maintenance and growth. This study investigated the flux of carbon from PEPC and direct Rubisco fixation to different leaf carbon pools and to phloem sap over the diurnal cycle. Concentrations and carbon isotope compositions of starch, soluble sugars, and organic acids were determined in leaves and phloem exudates of Kalanchoë daigremontiana Hamet et Perr., and related to CO(2) fixation by PEPC and Rubisco. Three types of leaf carbon pools could be distinguished. (i) Starch and malate pools were dominant and showed a pattern of reciprocal mobilization and accumulation (85/54 and 13/48 mg C g(-1) DW, respective, at the beginning/end of phase I). The carbon isotope composition of these pools was compatible with predominant PEPC fixation (delta(13)C values of -13 and -11 per thousand for starch and malate compared to -11 per thousand of PEPC fixed carbon). (ii) Isotopic composition (-17 per thousand and -14 per thousand) and concentration of glucose and fructose (2 and 3 mg C g(-1) DW, respectively) were not affected by diurnal metabolism, suggesting a low turnover. (iii) Sucrose (1-3 mg C g(-1) DW), in contrast, exhibited large diurnal changes in delta(13)C values (from -17 per thousand in the evening to -12 per thousand in the morning), which were not matched by net changes in sucrose concentration. This suggests a high sucrose turnover, fed by nocturnal starch degradation and direct Rubisco fixation during the day. A detailed dissection of the carbon fixation and mobilization pattern in K. daigremontiana revealed that direct fixation of Rubisco during the light accounted for 30% of phloem sucrose, but only 15% of fixed carbon, indicating that carbon from direct Rubisco fixation was preferentially used for leaf export.
Figures
References
-
- Borland AM. A model for the partitioning of photosynthetically fixed carbon during the C3-CAM transition in Sedum telephium. New Phytologist. 1996;134:433–444.
-
- Borland AM, Dodd AN. Carbohydrate partitioning in Crassulacean acid metabolism plants: reconciling potential conflicts of interest. Functional Plant Biology. 2002;29:707–716. - PubMed
-
- Borland AM, Griffiths H. Variations in the phases of Crassulacean acid metabolism and regulation of carboxylation patterns determined by carbon-isotope-discrimination techniques. Ecological Studies. 1996;114:230–249.
-
- Borland AM, Griffiths H, Broadmeadow MSJ, Fordham MC, Maxwell C. Carbon-isotope composition of biochemical fractions and the regulation of carbon balance in leaves of the C3-Crassulacean acid metabolism intermediate Clusia minor L growing in Trinidad. Plant Physiology. 1994;106:493–501. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
