

Increased insertion of glutamate receptor 2-lacking alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors at hippocampal synapses upon repeated morphine administration
- PMID: 20159947
- PMCID: PMC2872965
- DOI: 10.1124/mol.109.060301
Increased insertion of glutamate receptor 2-lacking alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors at hippocampal synapses upon repeated morphine administration
Abstract
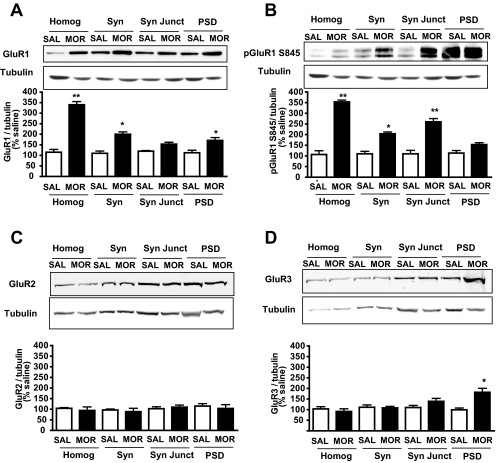
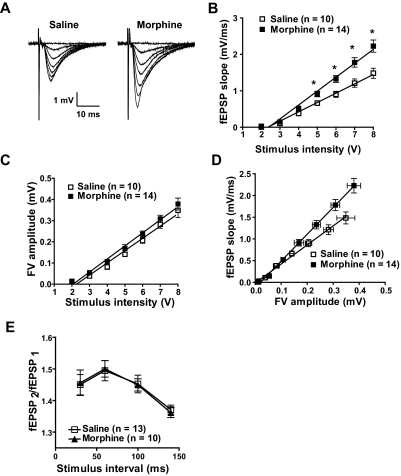
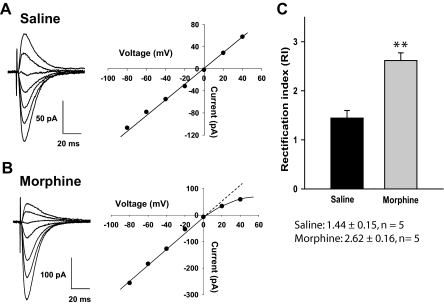
Evidence suggests that the long-term adaptations in the hippocampus after repeated drug treatment may parallel its role during memory formation. The neuroplasticity that subserves learning and memory is also believed to underlie addictive processes. We have reported previously that repeated morphine administration alters local distribution of endocytic proteins at hippocampal synapses, which could in turn affect expression of glutamate receptors. Glutamatergic systems, including alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptors (AMPARs), are believed to be involved in opiate-induced neuronal and behavioral plasticity, although the mechanisms underlying these effects are only beginning to be understood. The present study further examines the effects of repeated morphine administration on the expression and composition of AMPARs and the functional ramifications. Twelve hours after the last morphine injection, we observed an increased expression of AMPARs lacking glutamate receptor (GluR) 2 in hippocampal synaptic fractions. Immunoblotting studies show that 12 h after morphine treatment, GluR1 subunits are increased at the postsynaptic density (PSD) and at extrasynaptic sites, whereas GluR3 subunits are only increased at the PSD, and they show how this alters receptor subunit composition. In addition, we provide electrophysiological evidence that AMPARs are switched to Ca(2+)-permeable (GluR2-lacking) at the synapse 12 h after repeated morphine treatment, affecting the magnitude of long-term depression at hippocampal neurons. We propose that morphine-induced changes in glutamatergic synaptic transmission in the hippocampus may play an important role in the neuroadaptations induced by repeated morphine administration.
Figures




References
-
- Bellone C, Lüscher C. (2006) Cocaine triggered AMPA receptor redistribution is reversed in vivo by mGluR-dependent long-term depression. Nat Neurosci 9:636–641 - PubMed
-
- Chung HJ, Steinberg JP, Huganir RL, Linden DJ. (2003) Requirement of AMPA receptor GluR2 phosphorylation for cerebellar long-term depression. Science 300:1751–1755 - PubMed
-
- Colledge M, Dean RA, Scott GK, Langeberg LK, Huganir RL, Scott JD. (2000) Targeting of PKA to glutamate receptors through a MAGUK-AKAP complex. Neuron 27:107–119 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
