

Evaluation of the antigenic diversity of placenta-binding Plasmodium falciparum variants and the antibody repertoire among pregnant women
- PMID: 20160014
- PMCID: PMC2863515
- DOI: 10.1128/IAI.01365-09
Evaluation of the antigenic diversity of placenta-binding Plasmodium falciparum variants and the antibody repertoire among pregnant women
Abstract
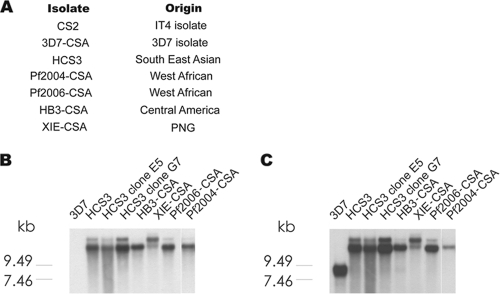
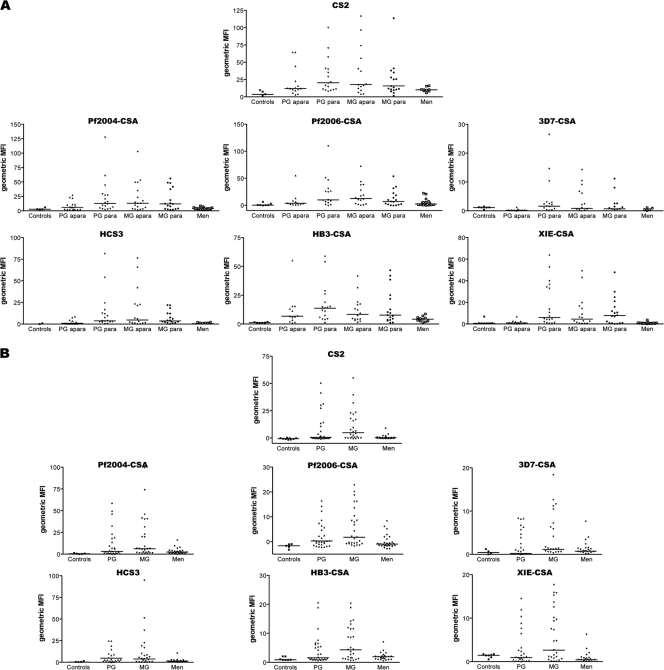
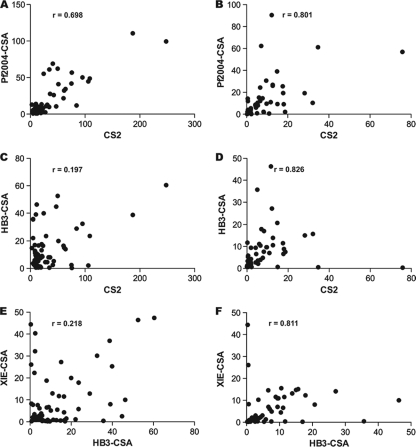
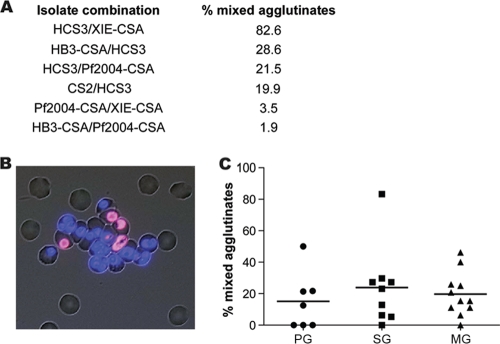
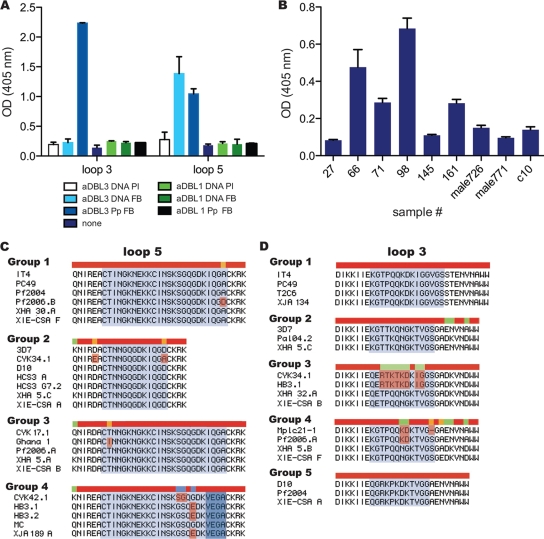
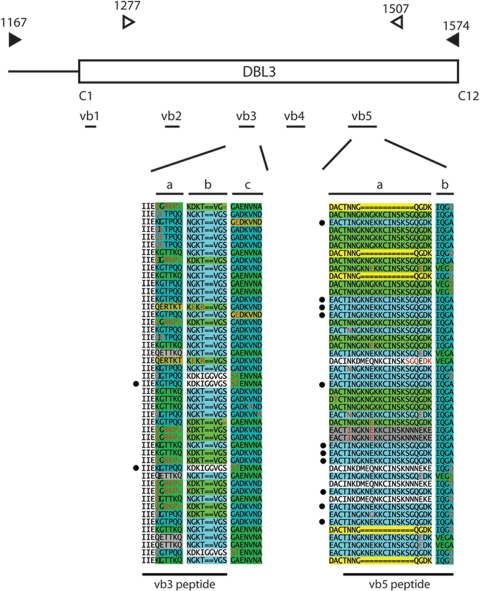
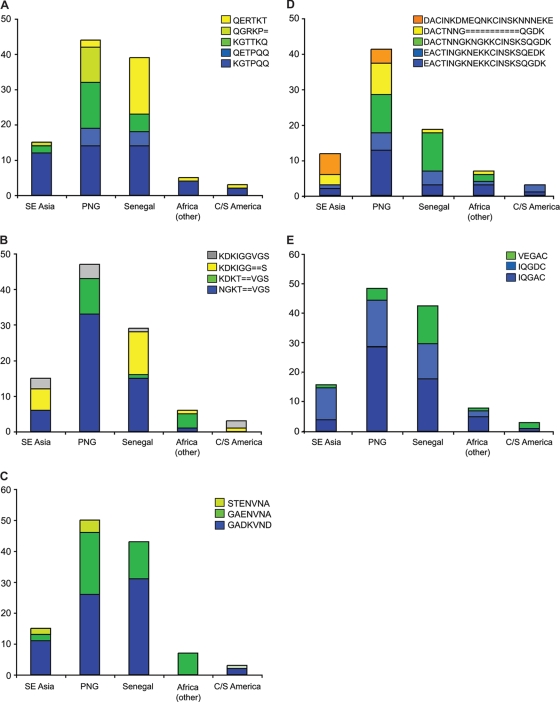
Pregnant women are infected by specific variants of Plasmodium falciparum that adhere and accumulate in the placenta. Using serological and molecular approaches, we assessed the global antigenic diversity of surface antigens expressed by placenta-binding isolates to better understand immunity to malaria in pregnancy and evolution of polymorphisms and to inform vaccine development. We found that placenta-binding isolates originating from all major regions where malaria occurs were commonly recognized by antibodies in different populations of pregnant women. There was substantial antigenic overlap and sharing of epitopes between isolates, including isolates from distant geographic locations, suggesting that there are limitations to antigenic diversity; however, differences between populations and isolates were also seen. Many women had cross-reactive antibodies and/or a broad repertoire of antibodies to different isolates. Studying VAR2CSA as the major antigen expressed by placenta-binding isolates, we identified antibody epitopes encoded by variable sequence blocks in the DBL3 domain. Analysis of global var2csa DBL3 sequences demonstrated that there was extensive sharing of variable blocks between Africa, Asia, Papua New Guinea, and Latin America, which likely contributes to the high level of antigenic overlap between different isolates. However, there was also evidence of geographic clustering of sequences and differences in VAR2CSA sequences between populations. The results indicate that there is limited antigenic diversity in placenta-binding isolates and may explain why immunity to malaria in pregnancy can be achieved after exposure during one pregnancy. Inclusion of a limited number of variants in a candidate vaccine may be sufficient for broad population coverage, but geographic considerations may also have to be included in vaccine design.
Figures
References
-
- Avril, M., B. R. Kulasekara, S. O. Gose, C. Rowe, M. Dahlbäck, P. E. Duffy, M. Fried, A. Salanti, L. Misher, D. L. Narum, and J. D. Smith. 2008. Evidence for globally shared, cross-reacting polymorphic epitopes in the pregnancy malaria vaccine candidate VAR2CSA. Infect. Immun. 74:1791-1800. - PMC - PubMed
-
- Awadalla, P. 2003. The evolutionary genomics of pathogen recombination. Nat. Rev. Genet. 4:50-60. - PubMed
-
- Barfod, L., N. L. Bernasconi, M. Dahlback, D. Jarrossay, P. H. Andersen, A. Salanti, M. F. Ofori, L. Turner, M. Resende, M. A. Nielsen, T. G. Theander, F. Sallusto, A. Lanzavecchia, and L. Hviid. 2007. Human pregnancy-associated malaria-specific B cells target polymorphic, conformational epitopes in VAR2CSA. Mol. Microbiol. 63:335-347. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
