

Protein Crystal Engineering of YpAC-IV using the Strategy of Excess Charge Reduction
- PMID: 20160955
- PMCID: PMC2758785
- DOI: 10.1021/cg9003142
Protein Crystal Engineering of YpAC-IV using the Strategy of Excess Charge Reduction
Abstract
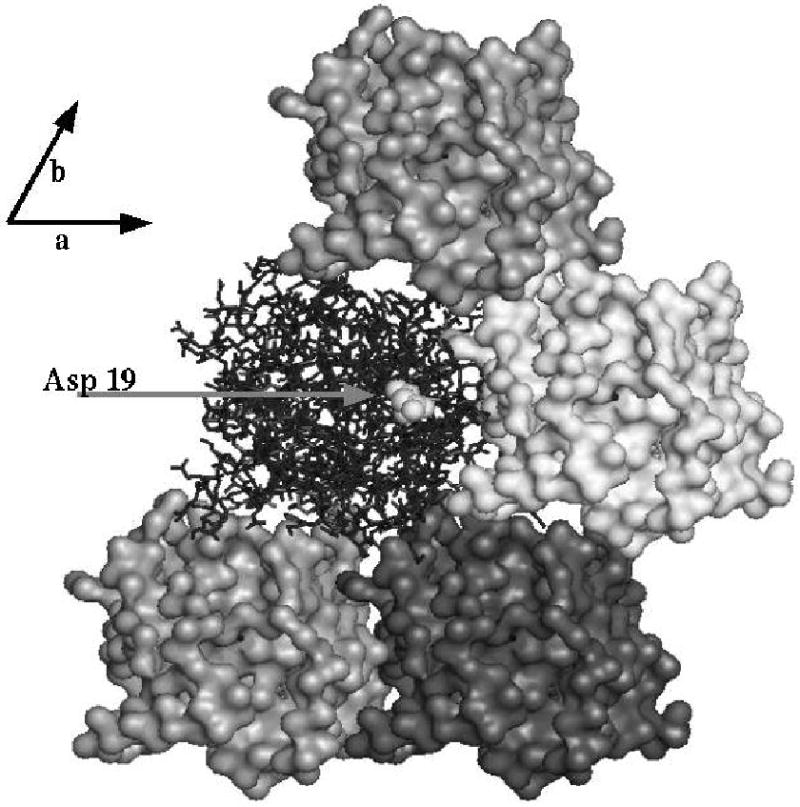
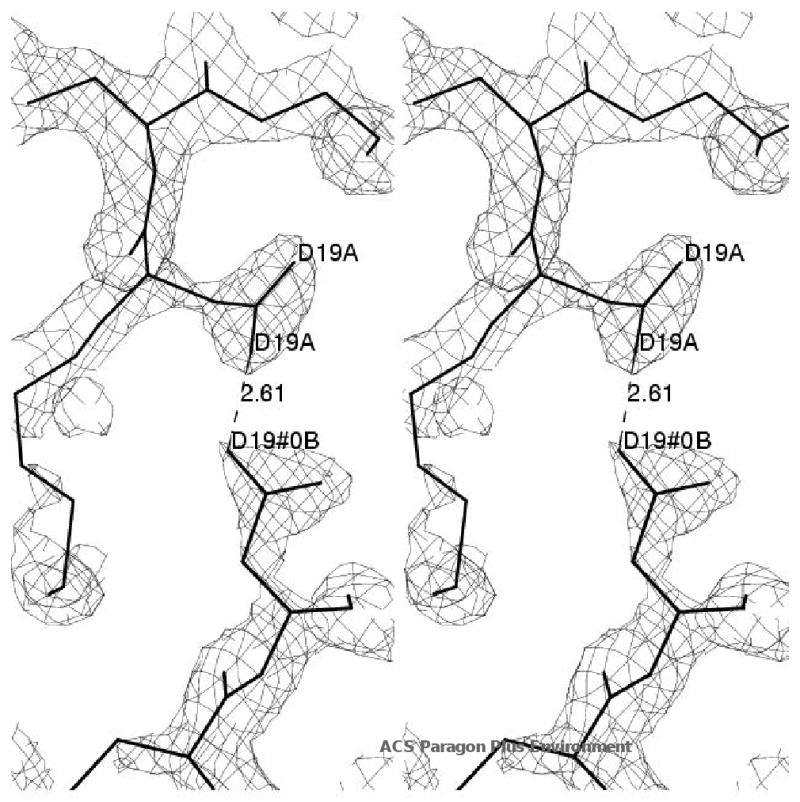
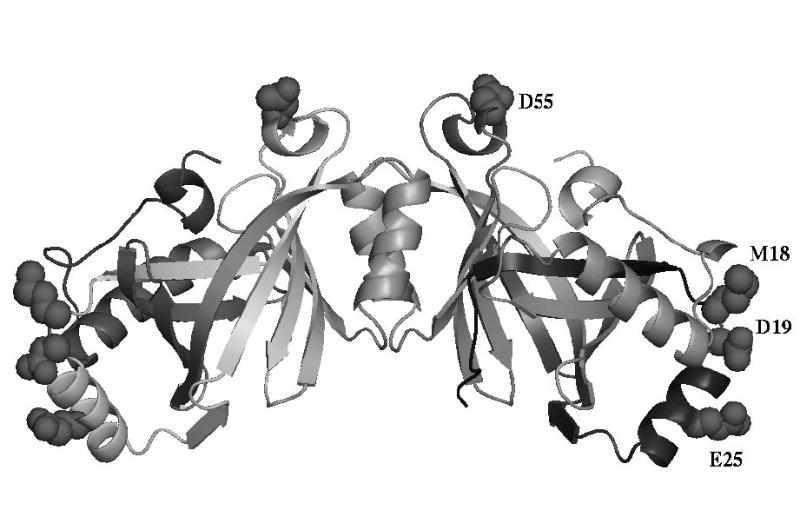
The class IV adenylyl cyclase from Yersinia pestis has been engineered by site-specific mutagenesis to facilitate crystallization at neutral pH. The wild-type enzyme crystallized only below pH 5, consistent with the observation of a carboxyl-carboxylate H bond in a crystal contact in the refined structure 2FJT. Based on that unliganded structure at 1.9 A resolution, two different approaches were tested with the goal of producing a higher-pH crystal needed for inhibitor complexation and mechanistic studies. In one approach, Asp 19, which forms the growth-limiting dicarboxyl contact in wild-type triclinic crystals, was modified to Ala and Asn in hopes of relieving the acid-dependence of that crystal form. In the other approach, wild-type residues Met 18, Glu 25, and Asp 55 were (individually) changed to lysine to reduce the protein's excess negative charge in hopes of enabling growth of new, higher-pH forms. These 3 sites were selected based on their high solvent exposure and lack of intraprotein interactions. The D19A and D19N mutants had reduced solubility and did not crystallize. The other 3 mutants all crystallized, producing several new forms at neutral pH. One of these forms, with the D55K mutant, enabled a product complex at 1.6 A resolution, structure 3GHX. This structure shows why the new crystal form required the mutation in order to grow at neutral pH. This approach could be useful in other cases where excess negative charge inhibits the crystallization of low-pI proteins.
Figures


Similar articles
-
Crystallization of the class IV adenylyl cyclase from Yersinia pestis.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006 Mar 1;62(Pt 3):200-4. doi: 10.1107/S1744309106002855. Epub 2006 Feb 10. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006. PMID: 16511301 Free PMC article.
-
Crystal structure analysis of a mutant Escherichia coli thioredoxin in which lysine 36 is replaced by glutamic acid.Biochemistry. 1993 May 18;32(19):5093-8. doi: 10.1021/bi00070a017. Biochemistry. 1993. PMID: 8098620
-
The impact of Glu-->Ala and Glu-->Asp mutations on the crystallization properties of RhoGDI: the structure of RhoGDI at 1.3 A resolution.Acta Crystallogr D Biol Crystallogr. 2002 Dec;58(Pt 12):1983-91. doi: 10.1107/s090744490201394x. Epub 2002 Nov 23. Acta Crystallogr D Biol Crystallogr. 2002. PMID: 12454455
-
Effects of buried charged groups on cysteine thiol ionization and reactivity in Escherichia coli thioredoxin: structural and functional characterization of mutants of Asp 26 and Lys 57.Biochemistry. 1997 Mar 4;36(9):2622-36. doi: 10.1021/bi961801a. Biochemistry. 1997. PMID: 9054569
-
Coupling of electron transfer to proton uptake at the Q(B) site of the bacterial reaction center: a perspective from FTIR difference spectroscopy.Biochim Biophys Acta. 2008 Oct;1777(10):1229-48. doi: 10.1016/j.bbabio.2008.06.012. Epub 2008 Jul 11. Biochim Biophys Acta. 2008. PMID: 18671937 Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous