

The phosphate transporter PiT1 (Slc20a1) revealed as a new essential gene for mouse liver development
- PMID: 20161774
- PMCID: PMC2818845
- DOI: 10.1371/journal.pone.0009148
The phosphate transporter PiT1 (Slc20a1) revealed as a new essential gene for mouse liver development
Abstract
Background: PiT1 (or SLC20a1) encodes a widely expressed plasma membrane protein functioning as a high-affinity Na(+)-phosphate (Pi) cotransporter. As such, PiT1 is often considered as a ubiquitous supplier of Pi for cellular needs regardless of the lack of experimental data. Although the importance of PiT1 in mineralizing processes have been demonstrated in vitro in osteoblasts, chondrocytes and vascular smooth muscle cells, in vivo evidence is missing.
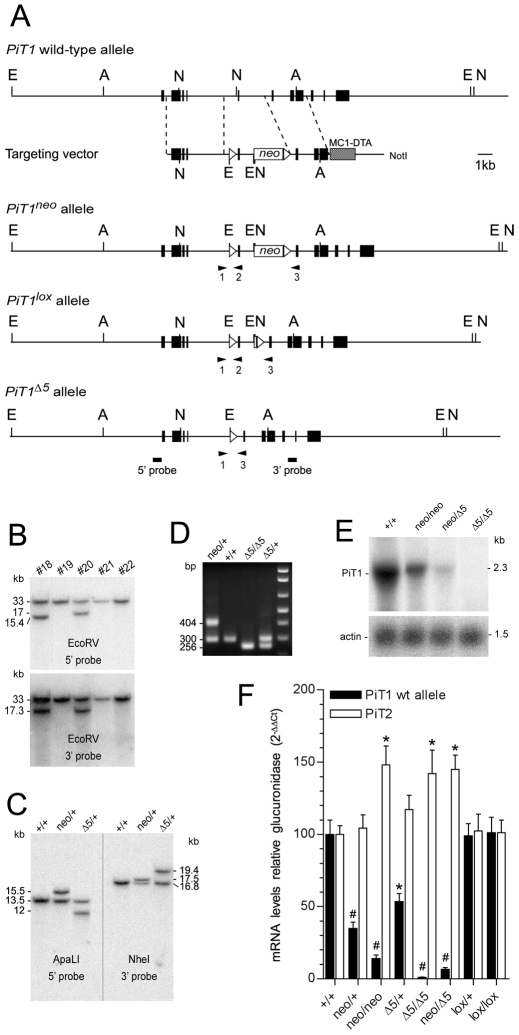
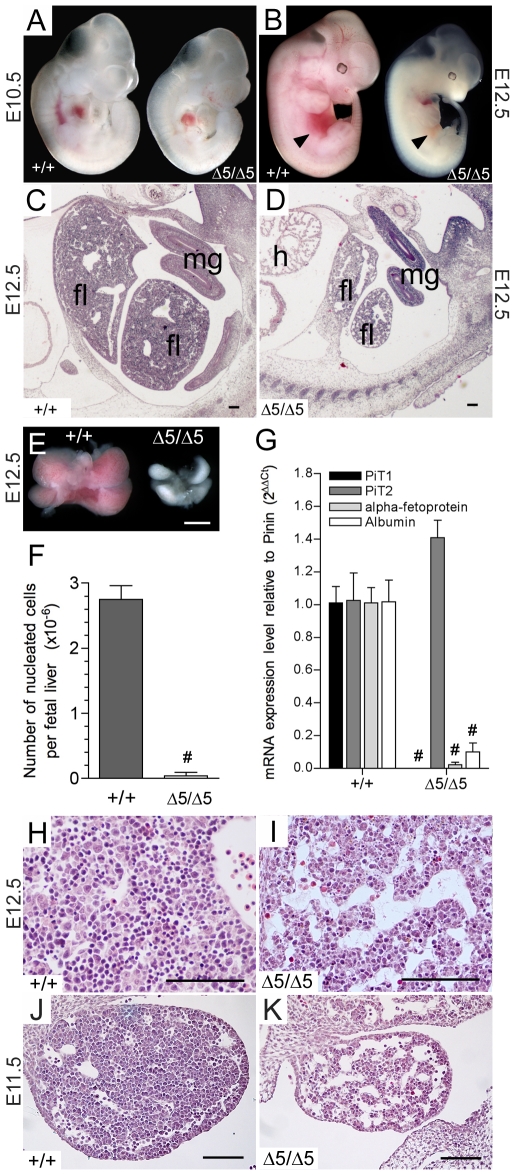
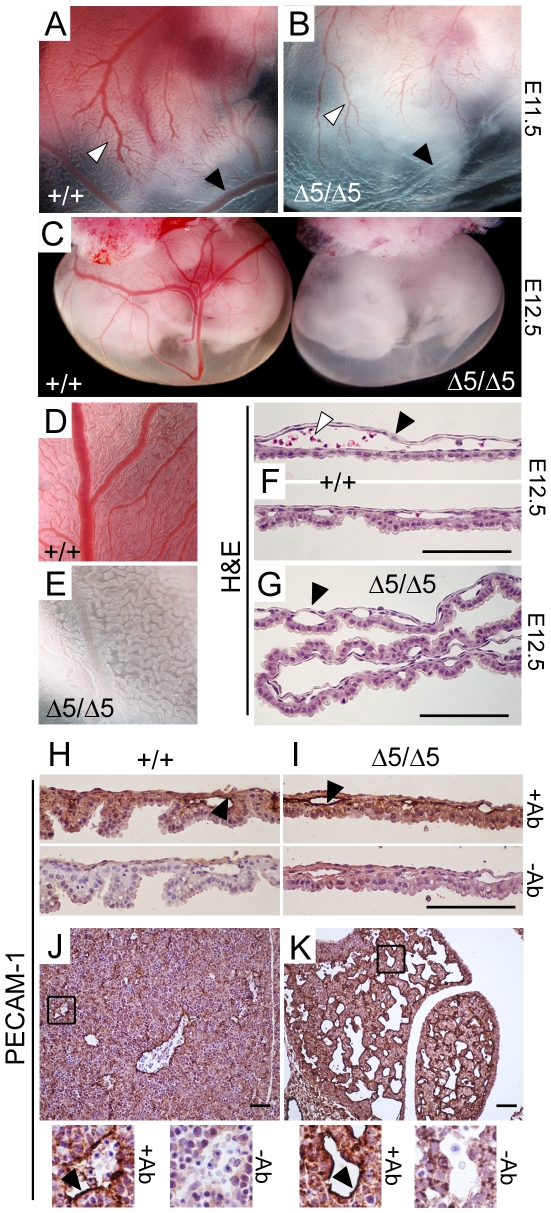
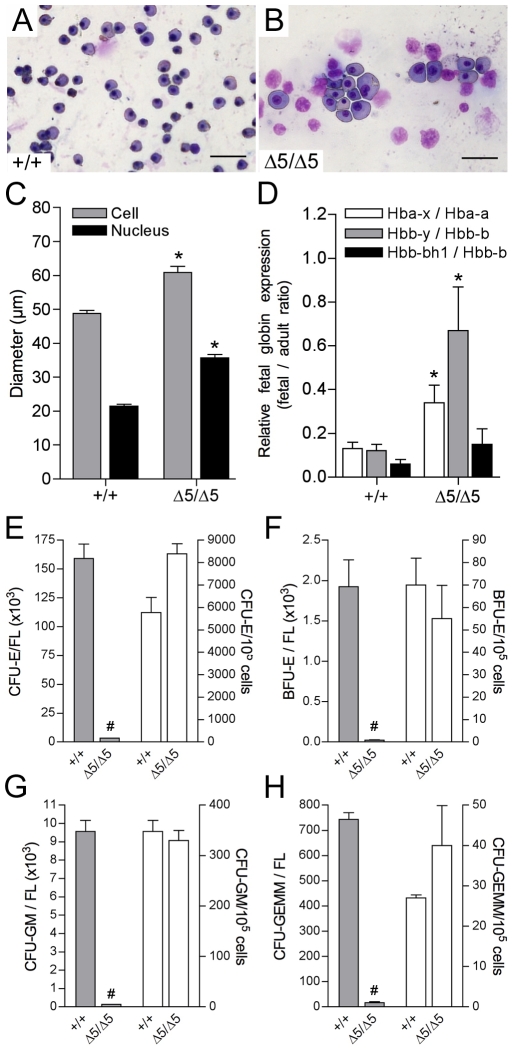
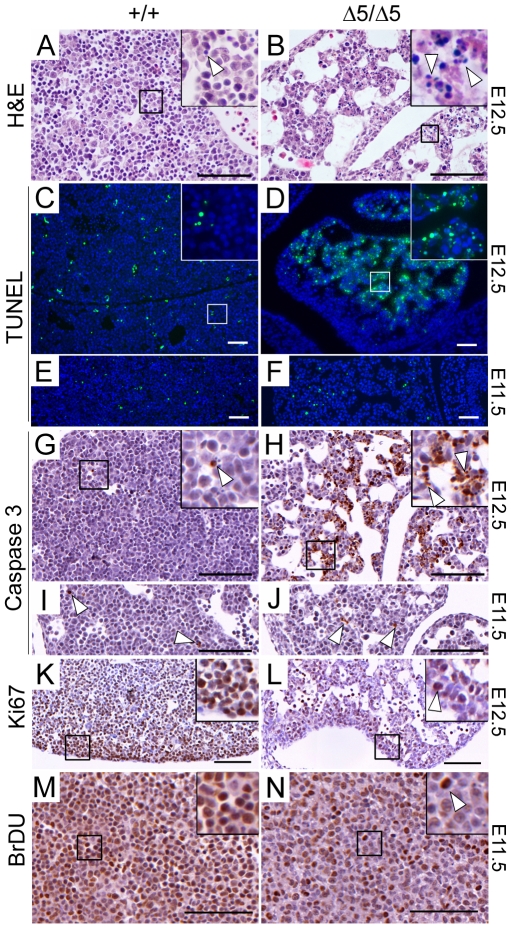
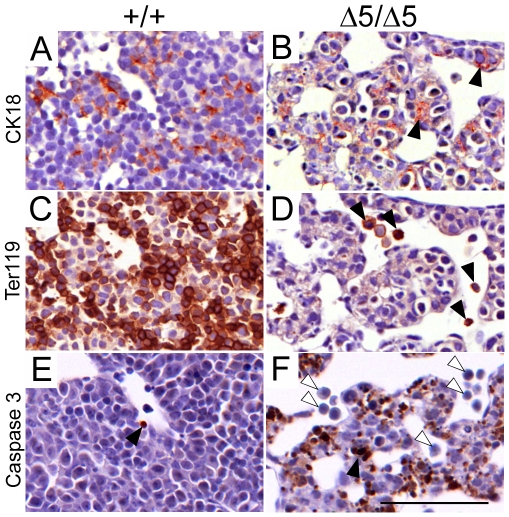
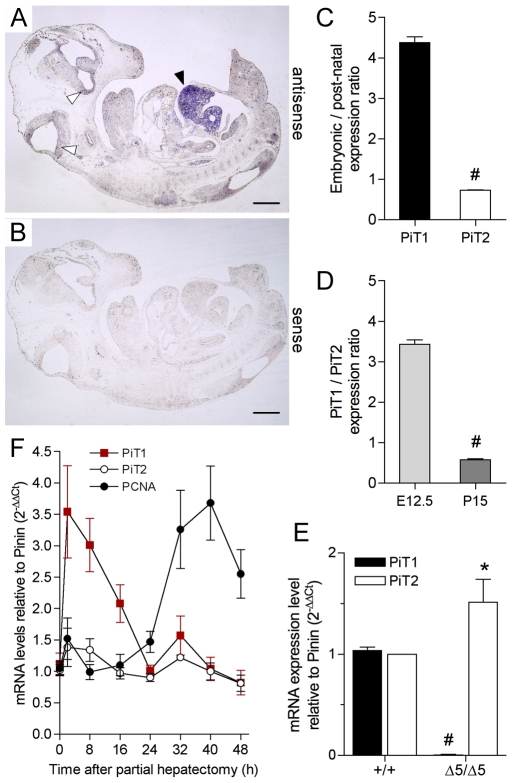
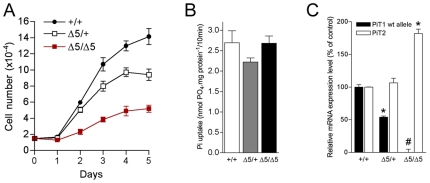
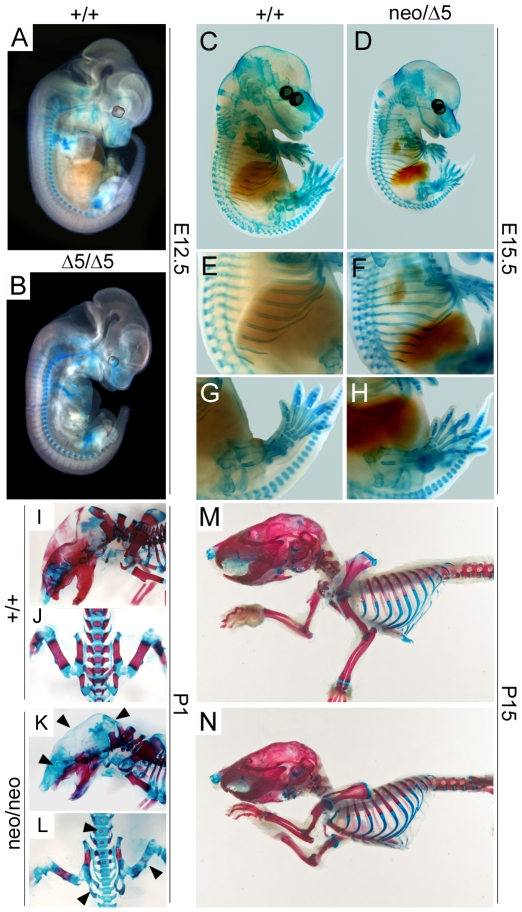
Methodology/principal findings: To determine the in vivo function of PiT1, we generated an allelic series of PiT1 mutations in mice by combination of wild-type, hypomorphic and null PiT1 alleles expressing from 100% to 0% of PiT1. In this report we show that complete deletion of PiT1 results in embryonic lethality at E12.5. PiT1-deficient embryos display severely hypoplastic fetal livers and subsequent reduced hematopoiesis resulting in embryonic death from anemia. We show that the anemia is not due to placental, yolk sac or vascular defects and that hematopoietic progenitors have no cell-autonomous defects in proliferation and differentiation. In contrast, mutant fetal livers display decreased proliferation and massive apoptosis. Animals carrying two copies of hypomorphic PiT1 alleles (resulting in 15% PiT1 expression comparing to wild-type animals) survive at birth but are growth-retarded and anemic. The combination of both hypomorphic and null alleles in heterozygous compounds results in late embryonic lethality (E14.5-E16.5) with phenotypic features intermediate between null and hypomorphic mice. In the three mouse lines generated we could not evidence defects in early skeleton formation.
Conclusion/significance: This work is the first to illustrate a specific in vivo role for PiT1 by uncovering it as being a critical gene for normal developmental liver growth.
Conflict of interest statement
Figures
References
-
- O'Hara B, Johann SV, Klinger HP, Blair DG, Rubinson H, et al. Characterization of a human gene conferring sensitivity to infection by gibbon ape leukemia virus. Cell Growth Differ. 1990;1:119–127. - PubMed
-
- Olah Z, Lehel C, Anderson WB, Eiden MV, Wilson CA. The cellular receptor for gibbon ape leukemia virus is a novel high affinity sodium-dependent phosphate transporter. J Biol Chem. 1994;269:25426–25431. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
