

Posttranslational modifications, localization, and protein interactions of optineurin, the product of a glaucoma gene
- PMID: 20161783
- PMCID: PMC2820081
- DOI: 10.1371/journal.pone.0009168
Posttranslational modifications, localization, and protein interactions of optineurin, the product of a glaucoma gene
Abstract
Background: Glaucoma is a major blinding disease. The most common form of this disease, primary open angle glaucoma (POAG), is genetically heterogeneous. One of the candidate genes, optineurin, is linked principally to normal tension glaucoma, a subtype of POAG. The present study was undertaken to illustrate the basic characteristics of optineurin.
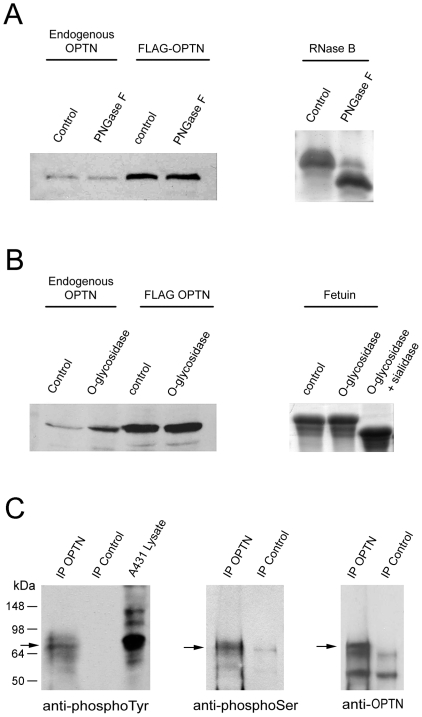
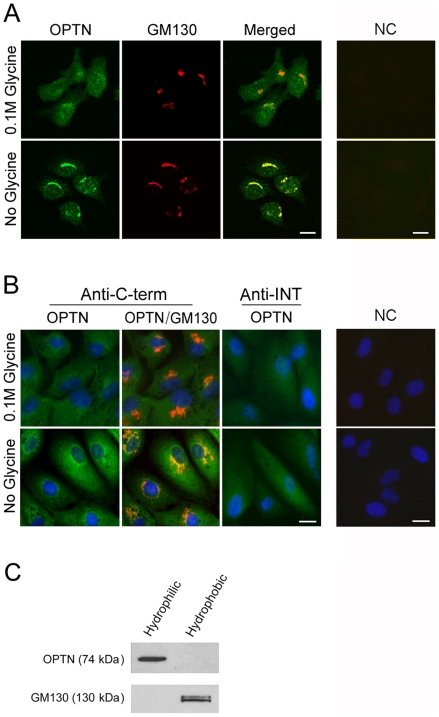
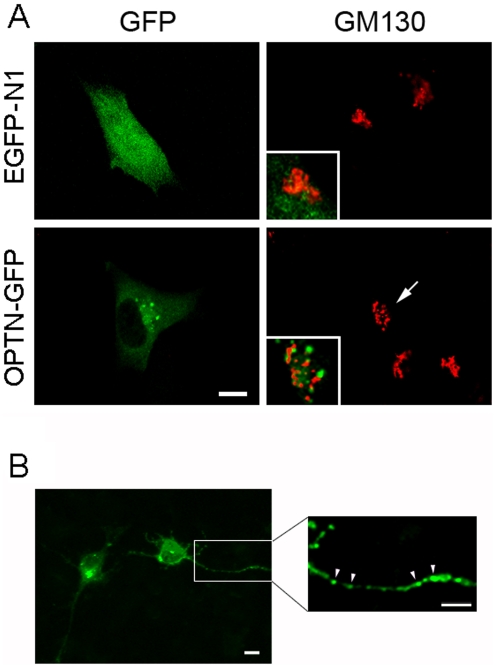
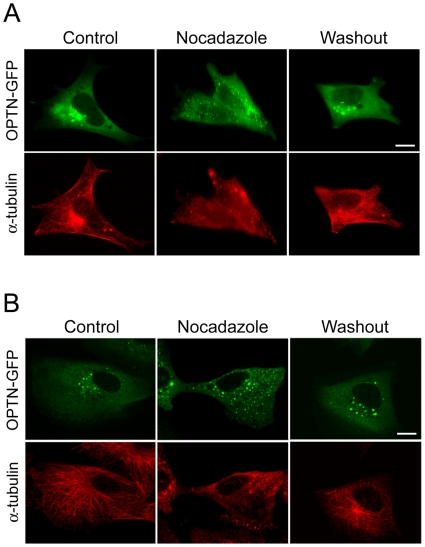
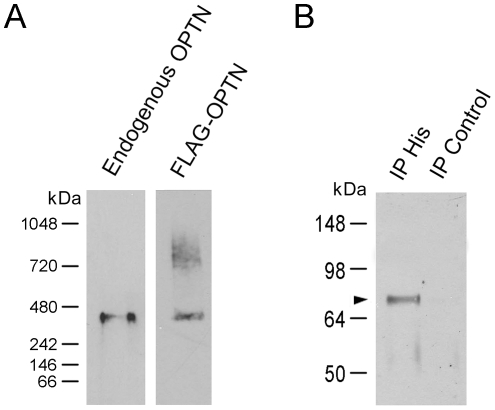
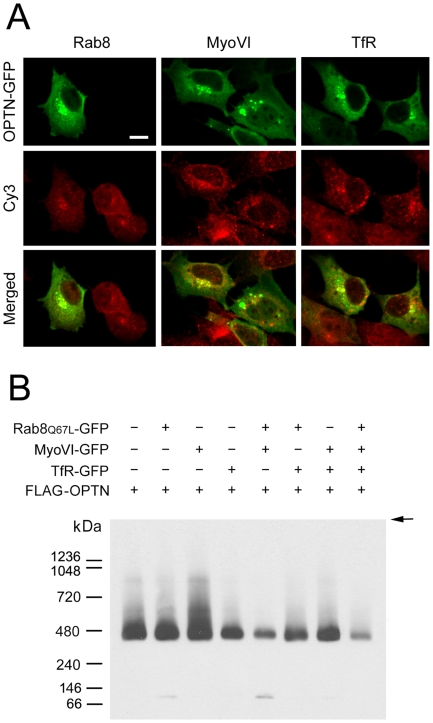
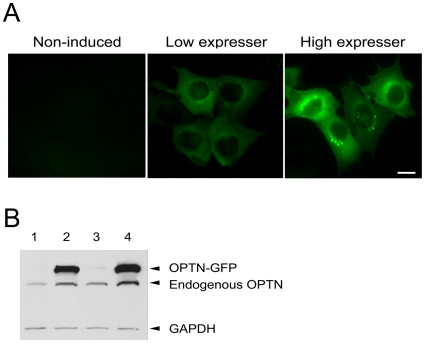
Methodology/principal findings: Lysates from rat retinal ganglion RGC5 cells were subjected to N- or O-deglycosylation or membrane protein extraction. The phosphorylation status was evaluated after immunoprecipitation. It was found that while phosphorylated, optineurin was neither N- nor O-glycosylated, and was by itself not a membrane protein. RGC5 and human retinal pigment epithelial cells were double stained with anti-optineurin and anti-GM130. The endogenous optineurin exhibited a diffuse, cytoplasmic distribution, but a population of the protein was associated with the Golgi apparatus. Turnover experiments showed that the endogenous optineurin was relatively short-lived, with a half-life of approximately 8 hours. Native blue gel electrophoresis revealed that the endogenous optineurin formed homohexamers. Optineurin also interacted with molecules including Rab8, myosin VI, and transferrin receptor to assemble into supermolecular complexes. When overexpressed, optineurin-green fluorescence protein (GFP) fusion protein formed punctate structures termed "foci" in the perinuclear region. Treatment of nocadazole resulted in dispersion of the optineurin foci. In addition, tetracycline-regulated optineurin-GFPs expressing RGC5 stable cell lines were established for the first time.
Conclusions/significance: The present study provides new information regarding basic characteristics of optineurin that are important for future efforts in defining precisely how optineurin functions normally and how mutations may result in pathology. The inducible optineurin-GFP-expressing cell lines are also anticipated to facilitate in-depth studies of optineurin. Furthermore, the demonstrations that optineurin is an aggregation-prone protein and that the foci formation is microtubule-dependent bear similarities to features documented in neurodegenerative diseases, supporting a neurodegenerative paradigm for glaucoma.
Conflict of interest statement
Figures

References
-
- Wiggs JL. Genetic etiologies of glaucoma. Arch Ophthalmol. 2007;125:30–37. - PubMed
-
- Fan BJ, Wang DY, Lam DS, Pang CP. Gene mapping for primary open angle glaucoma. Clin Biochem. 2006;39:249–258. - PubMed
-
- Rezaie T, Child A, Hitchings R, Brice G, Miller L, et al. Adult-onset primary open-angle glaucoma caused by mutations in optineurin. Science. 2002;295:1077–1079. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
