

The putative Na+/H+ antiporter of Vibrio cholerae, Vc-NhaP2, mediates the specific K+/H+ exchange in vivo
- PMID: 20163190
- PMCID: PMC2841436
- DOI: 10.1021/bi902173y
The putative Na+/H+ antiporter of Vibrio cholerae, Vc-NhaP2, mediates the specific K+/H+ exchange in vivo
Abstract
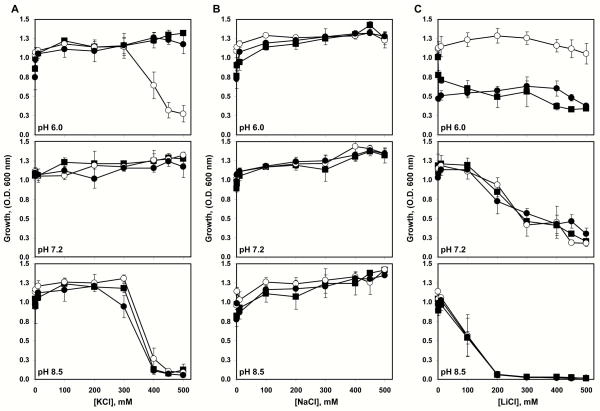
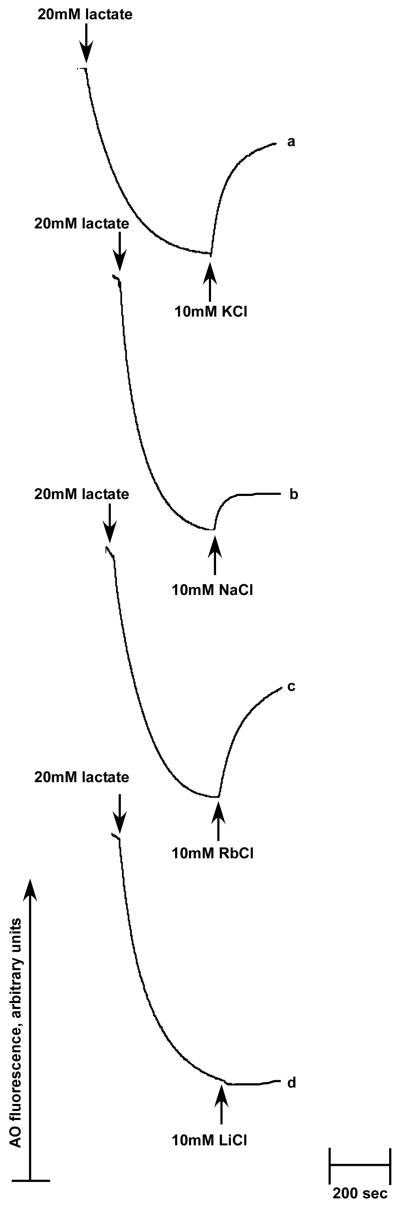
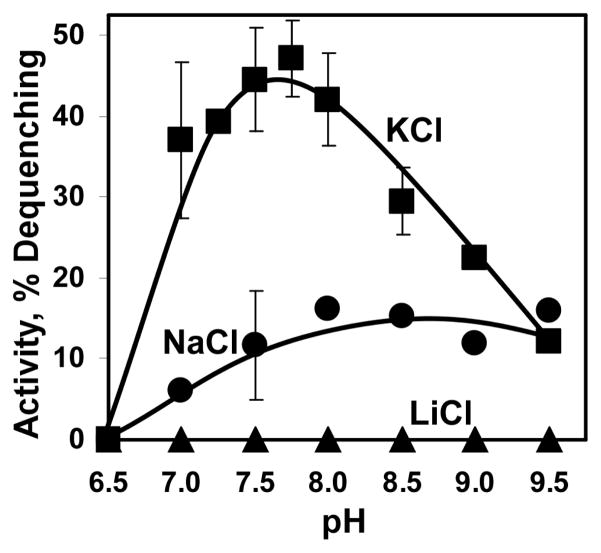
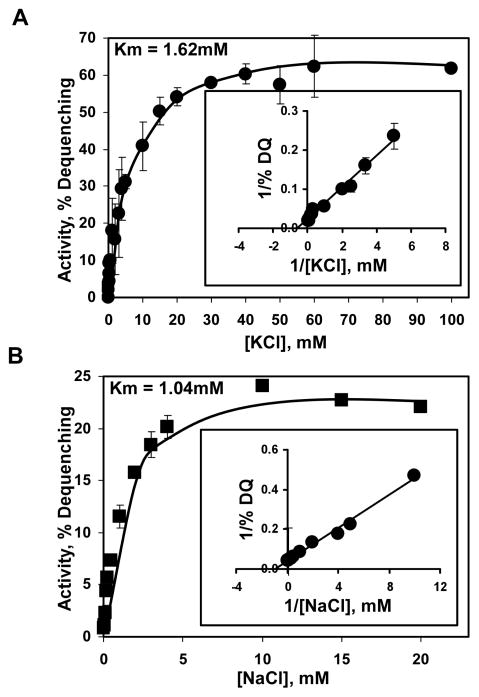
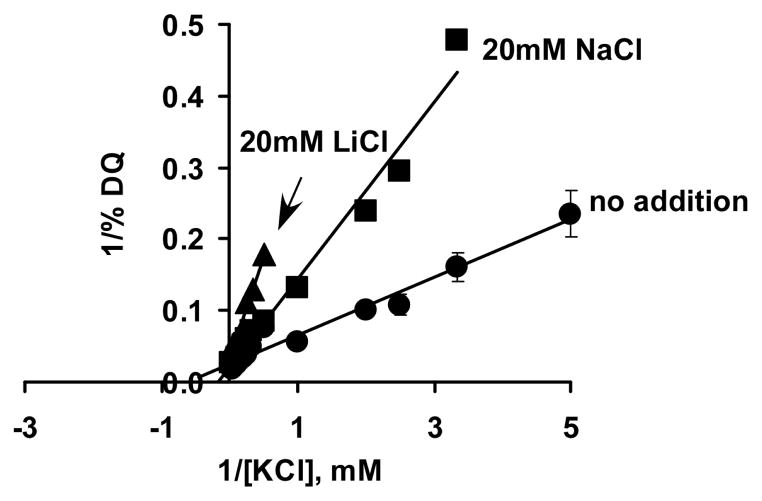
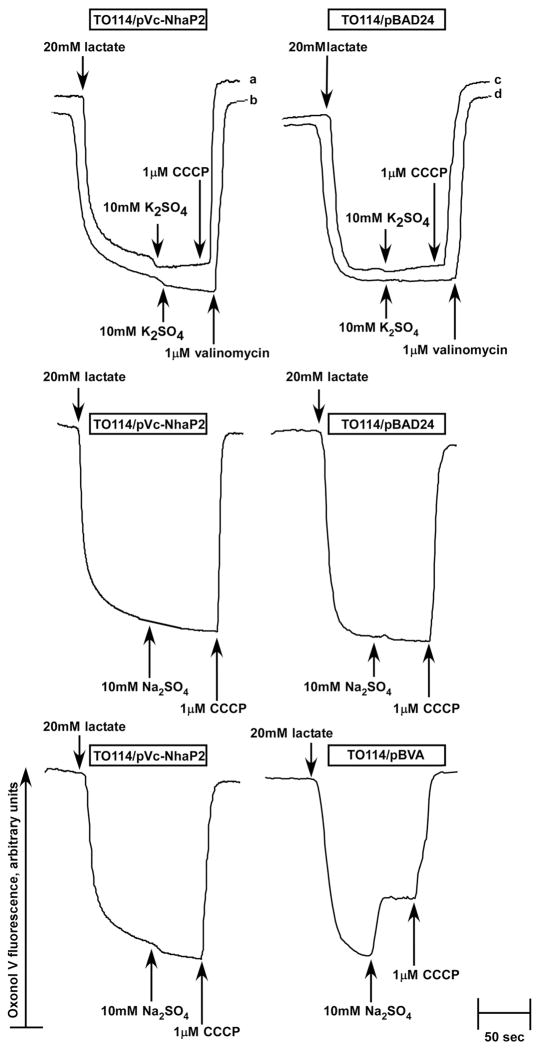
The existence of bacterial K(+)/H(+) antiporters that prevent the overaccumulation of potassium in the cytoplasm was predicted by Peter Mitchell almost 50 years ago. The importance of K(+)/H(+) antiport for bacterial physiology is widely recognized, but its molecular mechanisms remain underinvestigated. Here, we demonstrate that a putative Na(+)/H(+) antiporter, Vc-NhaP2, protects cells of Vibrio cholerae growing at pH 6.0 from high concentrations of external K(+). Resistance of V. cholerae to Na(+) was found to be independent of Vc-NhaP2. When assayed in inside-out membrane vesicles derived from antiporter-deficient Escherichia coli, Vc-NhaP2 catalyzed the electroneutral K(+)(Rb(+))/H(+) exchange with a pH optimum of approximately 7.75 with an apparent K(m) for K(+) of 1.62 mM. In the absence of K(+), it exhibited Na(+)/H(+) antiport, albeit rather weakly. Interestingly, while Vc-NhaP2 cannot exchange Li(+) for protons, elimination of functional Vc-NhaP2 resulted in a significantly higher Li(+) resistance of V. cholerae cells growing at pH 6.0, suggesting the possibility of Vc-NhaP2-mediated Li(+)/K(+) antiport. The peculiar cation specificity of Vc-NhaP2 and the presence of its two additional paralogues in the same genome make this transporter an attractive model for detailed analysis of the structural determinants of the substrate specificity in alkali cation exchangers.
Figures
Similar articles
-
Physiological, Structural, and Functional Analysis of the Paralogous Cation-Proton Antiporters of NhaP Type from Vibrio cholerae.Int J Mol Sci. 2019 May 25;20(10):2572. doi: 10.3390/ijms20102572. Int J Mol Sci. 2019. PMID: 31130620 Free PMC article. Review.
-
The C-terminal cytoplasmic portion of the NhaP2 cation-proton antiporter from Vibrio cholerae affects its activity and substrate affinity.Mol Cell Biochem. 2014 Apr;389(1-2):51-8. doi: 10.1007/s11010-013-1926-x. Epub 2013 Dec 18. Mol Cell Biochem. 2014. PMID: 24347178
-
Pharmacophore-based screening and modification of amiloride analogs for targeting the NhaP-type cation-proton antiporter in Vibrio cholerae.Can J Microbiol. 2021 Nov;67(11):835-849. doi: 10.1139/cjm-2021-0074. Epub 2021 Jul 5. Can J Microbiol. 2021. PMID: 34224663
-
A pathway leading to a cation-binding pocket determines the selectivity of the NhaP2 antiporter in Vibrio cholerae1.Biochem Cell Biol. 2019 Jun;97(3):307-314. doi: 10.1139/bcb-2018-0146. Epub 2018 Jul 16. Biochem Cell Biol. 2019. PMID: 30011386
-
Insights into the biochemistry of the ubiquitous NhaP family of cation/H+ antiporters.Biochem Cell Biol. 2011 Apr;89(2):130-7. doi: 10.1139/o10-149. Biochem Cell Biol. 2011. PMID: 21455265 Review.
Cited by
-
Effects of chromosomal deletion of the operon encoding the multiple resistance and pH-related antiporter in Vibrio cholerae.Microbiology (Reading). 2016 Dec;162(12):2147-2158. doi: 10.1099/mic.0.000384. Epub 2016 Oct 24. Microbiology (Reading). 2016. PMID: 27902431 Free PMC article.
-
Physiological, Structural, and Functional Analysis of the Paralogous Cation-Proton Antiporters of NhaP Type from Vibrio cholerae.Int J Mol Sci. 2019 May 25;20(10):2572. doi: 10.3390/ijms20102572. Int J Mol Sci. 2019. PMID: 31130620 Free PMC article. Review.
-
Geochemical and metagenomics study of a metal-rich, green-turquoise-coloured stream in the southern Swiss Alps.PLoS One. 2021 Mar 30;16(3):e0248877. doi: 10.1371/journal.pone.0248877. eCollection 2021. PLoS One. 2021. PMID: 33784327 Free PMC article.
-
Na+/H+ antiport is essential for Yersinia pestis virulence.Infect Immun. 2013 Sep;81(9):3163-72. doi: 10.1128/IAI.00071-13. Epub 2013 Jun 17. Infect Immun. 2013. PMID: 23774602 Free PMC article.
-
NhaP1 is a K+(Na+)/H+ antiporter required for growth and internal pH homeostasis of Vibrio cholerae at low extracellular pH.Microbiology (Reading). 2012 Apr;158(Pt 4):1094-1105. doi: 10.1099/mic.0.056119-0. Epub 2012 Jan 12. Microbiology (Reading). 2012. PMID: 22241048 Free PMC article.
References
-
- Epstein W. The roles and regulation of potassium in bacteria. Progress in Nucleic Acid Research. 2003;75:293–320. - PubMed
-
- Putnoky P, Kereszt A, Nakamura T, Endre G, Grosskopf E, Kiss P, Kondorosi A. The pha gene cluster of Rhizobium meliloti involved in pH adaptation and symbiosis encodes a novel type of K+ efflux system. Mol Microbiol. 1998;28:1091–101. - PubMed
-
- Benito B, Garciadeblás B, Rodriquez-Navarro A. Potassium- or sodium-efflux ATPase, a key enzyme in the evolution of fungi. Microbiology. 2002;148:933–941. - PubMed
-
- Radchenko MV, Waditee R, Oshimi S, Fukuhara M, Takabe T, Nakamura T. Cloning, functional expression and primary characterization of Vibrio parahaemolyticus K+/H+ antiporter genes in Escherichia coli. Mol Microbiol. 2006;59:651–663. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
