

Antibody isotype analysis of malaria-nematode co-infection: problems and solutions associated with cross-reactivity
- PMID: 20163714
- PMCID: PMC2838755
- DOI: 10.1186/1471-2172-11-6
Antibody isotype analysis of malaria-nematode co-infection: problems and solutions associated with cross-reactivity
Abstract
Background: Antibody isotype responses can be useful as indicators of immune bias during infection. In studies of parasite co-infection however, interpretation of immune bias is complicated by the occurrence of cross-reactive antibodies. To confidently attribute shifts in immune bias to the presence of a co-infecting parasite, we suggest practical approaches to account for antibody cross-reactivity. The potential for cross-reactive antibodies to influence disease outcome is also discussed.
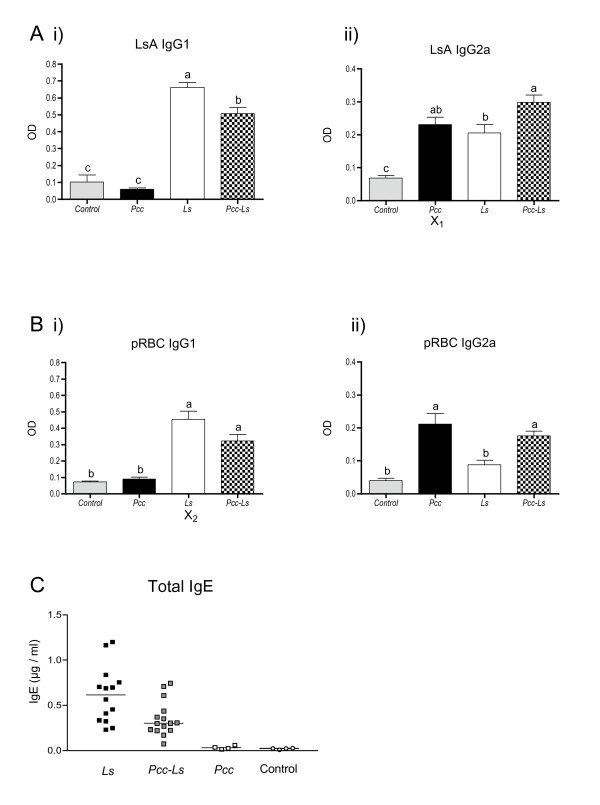
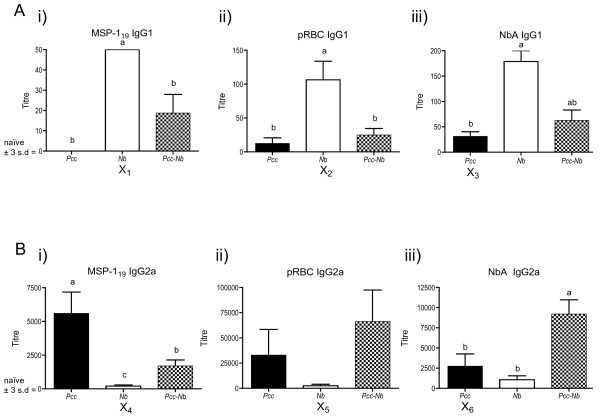
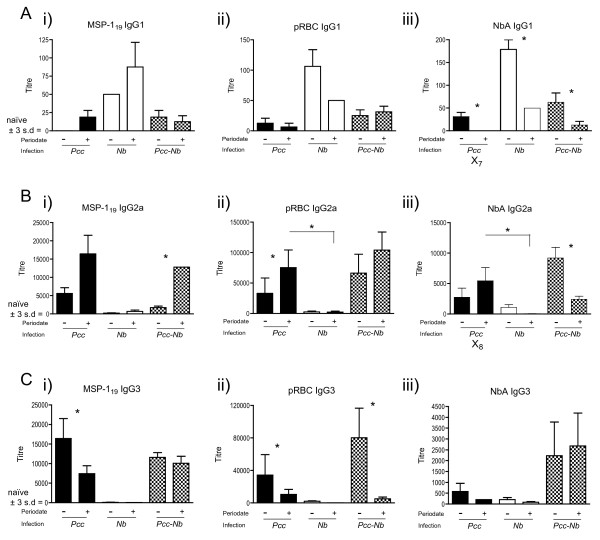
Results: Utilising two murine models of malaria-helminth co-infection we analysed antibody responses of mice singly- or co-infected with Plasmodium chabaudi chabaudi and Nippostrongylus brasiliensis or Litomosoides sigmodontis. We observed cross-reactive antibody responses that recognised antigens from both pathogens irrespective of whether crude parasite antigen preparations or purified recombinant proteins were used in ELISA. These responses were not apparent in control mice. The relative strength of cross-reactive versus antigen-specific responses was determined by calculating antibody titre. In addition, we analysed antibody binding to periodate-treated antigens, to distinguish responses targeted to protein versus carbohydrate moieties. Periodate treatment affected both antigen-specific and cross-reactive responses. For example, malaria-induced cross-reactive IgG1 responses were found to target the carbohydrate component of the helminth antigen, as they were not detected following periodate treatment. Interestingly, periodate treatment of recombinant malaria antigen Merozoite Surface Protein-119 (MSP-119) resulted in increased detection of antigen-specific IgG2a responses in malaria-infected mice. This suggests that glycosylation may have been masking protein epitopes and that periodate-treated MSP-119 may more closely reflect the natural non-glycosylated antigen seen during infection.
Conclusions: In order to utilize antibody isotypes as a measure of immune bias during co-infection studies, it is important to dissect antigen-specific from cross-reactive antibody responses. Calculating antibody titre, rather than using a single dilution of serum, as a measure of the relative strength of the response, largely accomplished this. Elimination of the carbohydrate moiety of an antigen that can often be the target of cross-reactive antibodies also proved useful.
Figures
Similar articles
-
Increased exposure to Plasmodium chabaudi antigens sustains cross-reactivity and avidity of antibodies binding Nippostrongylus brasiliensis: dissecting cross-phylum cross-reactivity in a rodent model.Parasitology. 2015 Dec;142(14):1703-14. doi: 10.1017/S0031182015001390. Epub 2015 Oct 22. Parasitology. 2015. PMID: 26490634
-
Plasmodium chabaudi limits early Nippostrongylus brasiliensis-induced pulmonary immune activation and Th2 polarization in co-infected mice.BMC Immunol. 2009 Dec 1;10:60. doi: 10.1186/1471-2172-10-60. BMC Immunol. 2009. PMID: 19951425 Free PMC article.
-
Protection against Plasmodium chabaudi malaria induced by immunization with apical membrane antigen 1 and merozoite surface protein 1 in the absence of gamma interferon or interleukin-4.Infect Immun. 2004 Oct;72(10):5605-12. doi: 10.1128/IAI.72.10.5605-5612.2004. Infect Immun. 2004. PMID: 15385457 Free PMC article.
-
Malaria-specific antibody responses and parasite persistence after infection of mice with Plasmodium chabaudi chabaudi.Parasite Immunol. 2007 Sep;29(9):435-44. doi: 10.1111/j.1365-3024.2007.00960.x. Parasite Immunol. 2007. PMID: 17727567
-
Quantifying variation in the potential for antibody-mediated apparent competition among nine genotypes of the rodent malaria parasite Plasmodium chabaudi.Infect Genet Evol. 2013 Dec;20:270-5. doi: 10.1016/j.meegid.2013.09.013. Epub 2013 Sep 18. Infect Genet Evol. 2013. PMID: 24056014 Free PMC article.
Cited by
-
Parasitic nematodes simultaneously suppress and benefit from coccidian coinfection in their natural mouse host.Parasitology. 2019 Jul;146(8):1096-1106. doi: 10.1017/S0031182019000192. Epub 2019 Mar 27. Parasitology. 2019. PMID: 30915927 Free PMC article.
-
Deletion of parasite immune modulatory sequences combined with immune activating signals enhances vaccine mediated protection against filarial nematodes.PLoS Negl Trop Dis. 2012;6(12):e1968. doi: 10.1371/journal.pntd.0001968. Epub 2012 Dec 27. PLoS Negl Trop Dis. 2012. PMID: 23301106 Free PMC article.
-
Malaria endemicity and co-infection with tissue-dwelling parasites in Sub-Saharan Africa: a review.Infect Dis Poverty. 2015 Aug 29;4:35. doi: 10.1186/s40249-015-0070-0. Infect Dis Poverty. 2015. PMID: 26377900 Free PMC article. Review.
-
Serology- and Blood-PCR-Based Screening for Schistosomiasis in Pregnant Women in Madagascar-A Cross-Sectional Study and Test Comparison Approach.Pathogens. 2021 Jun 8;10(6):722. doi: 10.3390/pathogens10060722. Pathogens. 2021. PMID: 34201231 Free PMC article.
-
Influenza virus-like particle vaccine containing both apical membrane antigen 1 and microneme-associated antigen proteins of Plasmodium berghei confers protection in mice.BMC Immunol. 2022 Apr 25;23(1):21. doi: 10.1186/s12865-022-00494-4. BMC Immunol. 2022. PMID: 35468726 Free PMC article.
References
-
- Yatich NJ, Yi J, Agbenyega T, Turpin A, Rayner JC, Stiles JK, Ellis WO, Funkhouser E, Ehiri JE, Williams JH, Jolly PE. Malaria and intestinal helminth co-infection among pregnant women in Ghana: prevalence and risk factors. Am J Trop Med Hyg. 2009;80:896–901. - PubMed
-
- NJ Faye B, Tine RC, Lô AC, Gaye O. [Interaction between malaria and intestinal helminthiasis in Senegal: influence of the carriage of intestinal parasites on the intensity of the malaria infection] Bulletin de la Societe pathologie exotique. 2008;101:391–394. doi: 10.3185/pathexo3189. - DOI - PubMed
-
- Piessens WF, Hoffman SL, Ratiwayanto S, Piessens PW, Partono F, Kurniawan L, Marwoto HA. Opposing effects of filariasis and chronic malaria on immunoregulatory T lymphocytes. Diagn Immunol. 1983;1:257–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
