

Hantavirus nucleocapsid protein has distinct m7G cap- and RNA-binding sites
- PMID: 20164193
- PMCID: PMC2857014
- DOI: 10.1074/jbc.M110.102459
Hantavirus nucleocapsid protein has distinct m7G cap- and RNA-binding sites
Abstract
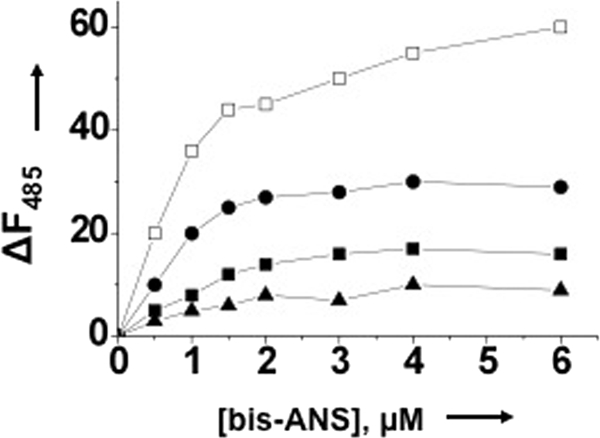
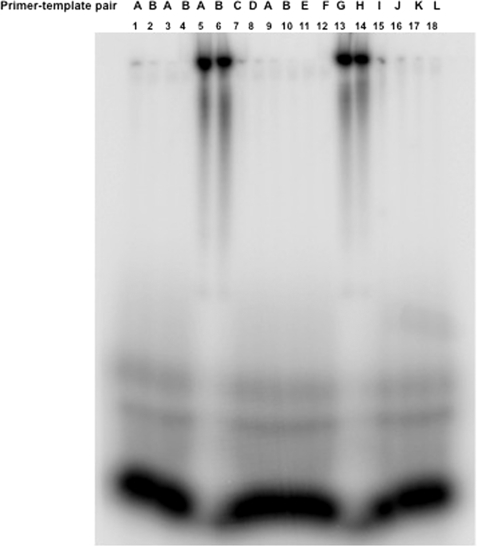
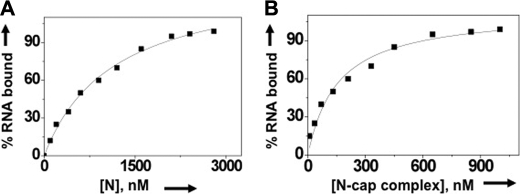
Hantaviruses, members of the Bunyaviridae family, are emerging category A pathogens that carry three negative stranded RNA molecules as their genome. Hantavirus nucleocapsid protein (N) is encoded by the smallest S segment genomic RNA (viral RNA). N specifically binds mRNA caps and requires four nucleotides adjacent to the cap for high affinity binding. We show that the N peptide has distinct cap- and RNA-binding sites that independently interact with mRNA cap and viral genomic RNA, respectively. In addition, N can simultaneously bind with both mRNA cap and vRNA. N undergoes distinct conformational changes after binding with either mRNA cap or vRNA or both mRNA cap and vRNA simultaneously. Hantavirus RNA-dependent RNA polymerase (RdRp) uses a capped RNA primer for transcription initiation. The capped RNA primer is generated from host cell mRNA by the cap-snatching mechanism and is supposed to anneal with the 3' terminus of vRNA template during transcription initiation by single G-C base pairing. We show that the capped RNA primer binds at the cap-binding site and induces a conformational change in N. The conformationally altered N with a capped primer loaded at the cap-binding site specifically binds the conserved 3' nine nucleotides of vRNA and assists the bound primer to anneal at the 3' terminus. We suggest that the cap-binding site of N, in conjunction with RdRp, plays a key role during the transcription and replication initiation of vRNA genome.
Figures
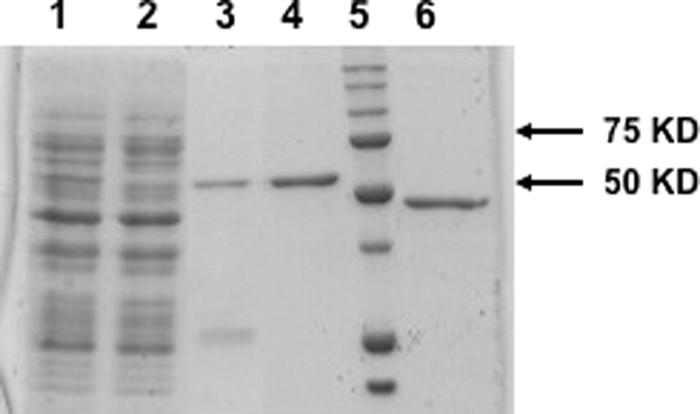
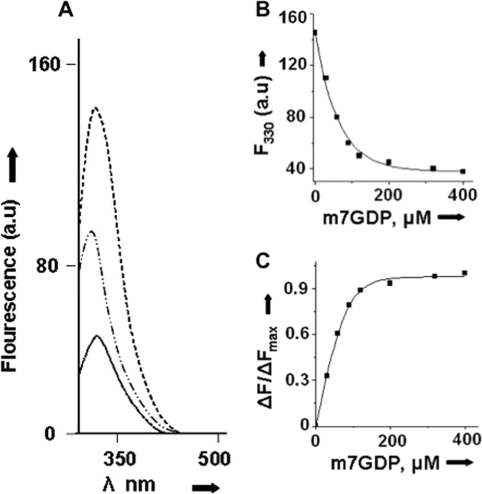
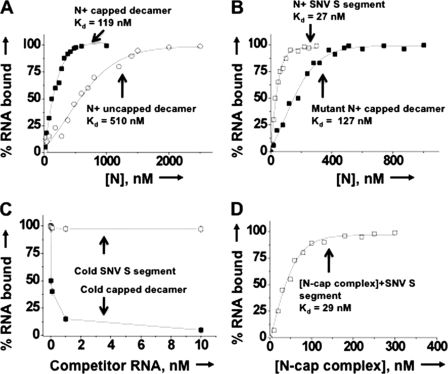

Similar articles
-
Signatures of host mRNA 5' terminus for efficient hantavirus cap snatching.J Virol. 2012 Sep;86(18):10173-85. doi: 10.1128/JVI.05560-11. Epub 2012 Jul 11. J Virol. 2012. PMID: 22787213 Free PMC article.
-
Hantavirus RdRp Requires a Host Cell Factor for Cap Snatching.J Virol. 2019 Feb 19;93(5):e02088-18. doi: 10.1128/JVI.02088-18. Print 2019 Mar 1. J Virol. 2019. PMID: 30541836 Free PMC article.
-
Interaction of hantavirus nucleocapsid protein with ribosomal protein S19.J Virol. 2010 Dec;84(23):12450-3. doi: 10.1128/JVI.01388-10. Epub 2010 Sep 15. J Virol. 2010. PMID: 20844026 Free PMC article.
-
How hantaviruses modulate cellular pathways for efficient replication?Front Biosci (Elite Ed). 2013 Jan 1;5(1):154-66. doi: 10.2741/e604. Front Biosci (Elite Ed). 2013. PMID: 23276978 Review.
-
Insight into Influenza: A Virus Cap-Snatching.Viruses. 2018 Nov 16;10(11):641. doi: 10.3390/v10110641. Viruses. 2018. PMID: 30453478 Free PMC article. Review.
Cited by
-
The triplet repeats of the Sin Nombre hantavirus 5' untranslated region are sufficient in cis for nucleocapsid-mediated translation initiation.J Virol. 2010 Sep;84(17):8937-44. doi: 10.1128/JVI.02720-09. Epub 2010 Jun 23. J Virol. 2010. PMID: 20573811 Free PMC article.
-
Crystal Structure of the Core Region of Hantavirus Nucleocapsid Protein Reveals the Mechanism for Ribonucleoprotein Complex Formation.J Virol. 2015 Nov 11;90(2):1048-61. doi: 10.1128/JVI.02523-15. Print 2016 Jan 15. J Virol. 2015. PMID: 26559827 Free PMC article.
-
Structural characterization of the full-length Hantaan virus polymerase.PLoS Pathog. 2024 Dec 9;20(12):e1012781. doi: 10.1371/journal.ppat.1012781. eCollection 2024 Dec. PLoS Pathog. 2024. PMID: 39652621 Free PMC article.
-
Genetic reassortment between high-virulent and low-virulent Dobrava-Belgrade virus strains.Virus Genes. 2010 Dec;41(3):319-28. doi: 10.1007/s11262-010-0523-2. Epub 2010 Aug 24. Virus Genes. 2010. PMID: 20734125
-
Ribosomal protein S19-binding domain provides insights into hantavirus nucleocapsid protein-mediated translation initiation mechanism.Biochem J. 2014 Nov 15;464(1):109-21. doi: 10.1042/BJ20140449. Biochem J. 2014. PMID: 25062117 Free PMC article.
References
-
- Schmaljohn C. M. (1996) Molecular Biology of Hantaviruses, pp. 1–20, Plenum Press, New York
-
- Schmaljohn C. S., Hooper J. W. (2001) in Fields Virology (Howley K. A. ed.) pp. 1581–1602, Lippencott, Williams, and Wilkins, Philadelphia:
-
- Schmaljohn C. S., Jonsson C. B. (2001) in Hantaviruses (Nichol S. A. ed) pp. 15–32, Springer-Verlag, Berlin
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
