

Interactive changes between macrophages and adipocytes
- PMID: 20164250
- PMCID: PMC2849320
- DOI: 10.1128/CVI.00494-09
Interactive changes between macrophages and adipocytes
Abstract
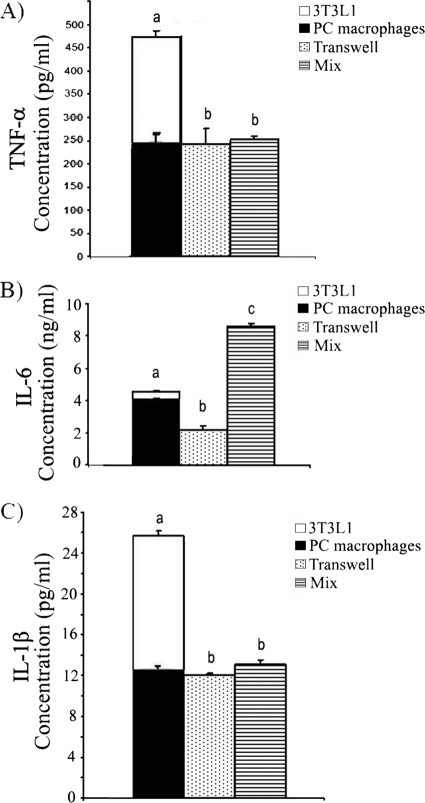
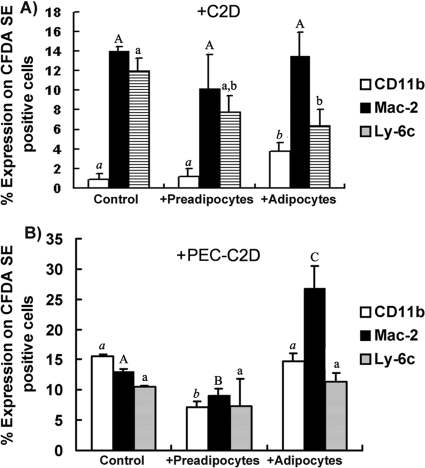
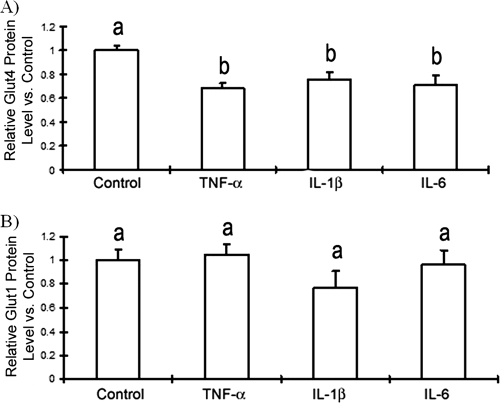
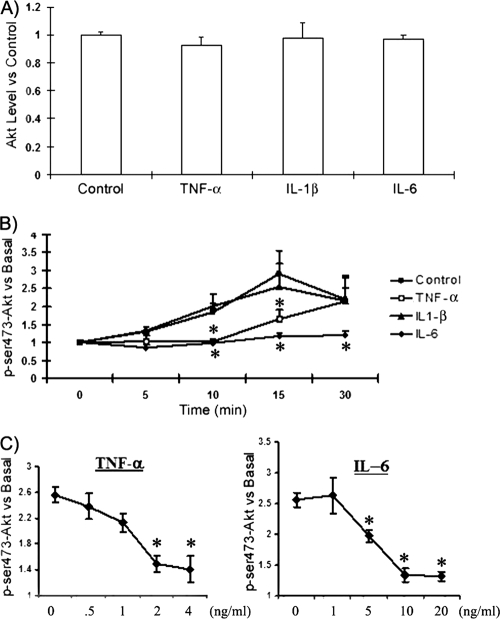
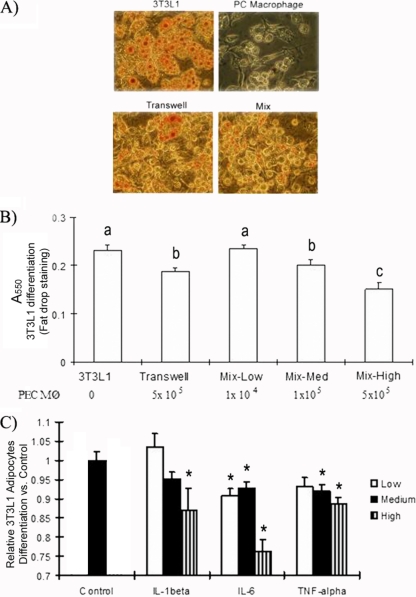
Obesity is associated with a proinflammatory state, with macrophage infiltration into adipose tissue. We tested the hypothesis that communication between macrophages and adipocytes affects insulin resistance by disrupting insulin-stimulated glucose transport, adipocyte differentiation, and macrophage function. To test this hypothesis, we cocultured 3T3-L1 adipocytes with C2D macrophages or primary peritoneal mouse macrophages and examined the impacts of macrophages and adipocytes on each other. Adipocytes and preadipocytes did not affect C2D macrophage TNF-alpha, IL-6, or IL-1beta transcript concentrations relative to those obtained when C2D macrophages were incubated alone. However, preadipocytes and adipocytes increased PEC-C2D macrophage IL-6 transcript levels, while preadipocytes inhibited IL-1beta transcript levels compared to those obtained when PEC-C2D macrophages were incubated in medium alone. We found that adipocyte coculture increased macrophage consumption of tumor necrosis factor alpha (TNF-alpha), interleukin 1beta (IL-1beta), and, in some cases, IL-6. C2D macrophages increasingly downregulated GLUT4 transcript levels in differentiated adipocytes. Recombinant TNF-alpha, IL-1beta, and IL-6 also downregulated GLUT4 transcript levels relative to those for the control. However, only IL-6 was inhibitory at concentrations detected in macrophage-adipocyte cocultures. IL-6 and TNF-alpha, but not IL-1beta, inhibited Akt phosphorylation within 15 min of insulin stimulation, but only IL-6 was inhibitory 30 min after stimulation. Lastly, we found that adipocyte differentiation was inhibited by macrophages or by recombinant TNF-alpha, IL-6, and IL-1beta, with IL-6 having the most impact. These data suggest that the interaction between macrophages and adipocytes is a complex process, and they support the hypothesis that the macrophage-adipocyte interaction affects insulin resistance by disrupting insulin-stimulated glucose transport, adipocyte differentiation, and macrophage function.
Figures
References
-
- Ailhaud, G. 2006. Adipose tissue as a secretory organ: from adipogenesis to the metabolic syndrome. C. R. Biol. 329:570-577. - PubMed
-
- Ajuwon, K. M., and M. E. Spurlock. 2005. Adiponectin inhibits LPS-induced NF-κB activation and IL-6 production and increases PPARγ2 expression in adipocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 288:R1220-R1225. - PubMed
-
- Beharka, A. A., J. W. Armstrong, and S. K. Chapes. 1998. Macrophage cell lines derived from major histocompatibility complex II-negative mice. In Vitro Cell. Dev. Biol. Anim. 34:499-507. - PubMed
-
- Blüher, M., M. Fasshauer, A. Tonjes, J. Kratzsch, M. R. Schon, and R. Paschke. 2005. Association of interleukin-6, C-reactive protein, interleukin-10 and adiponectin plasma concentrations with measures of obesity, insulin sensitivity and glucose metabolism. Exp. Clin. Endocrinol. Diabetes 113:534-537. - PubMed
-
- Carey, A. L., G. R. Steinberg, S. L. Macaulay, W. G. Thomas, A. G. Holmes, G. Ramm, O. Prelovsek, C. Hohnen-Behrens, M. J. Watt, D. E. James, B. E. Kemp, B. K. Pedersen, and M. A. Febbraio. 2006. Interleukin-6 increases insulin-stimulated glucose disposal in humans and glucose uptake and fatty acid oxidation in vitro via AMP-activated protein kinase. Diabetes 55:2688-2697. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
