

Differential regulation of behavioral tolerance to WIN55,212-2 by GASP1
- PMID: 20164830
- PMCID: PMC2953419
- DOI: 10.1038/npp.2010.6
Differential regulation of behavioral tolerance to WIN55,212-2 by GASP1
Abstract
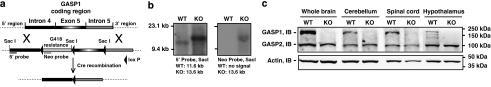
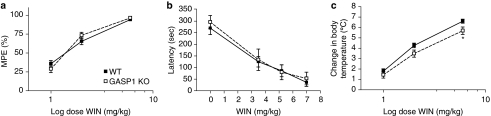
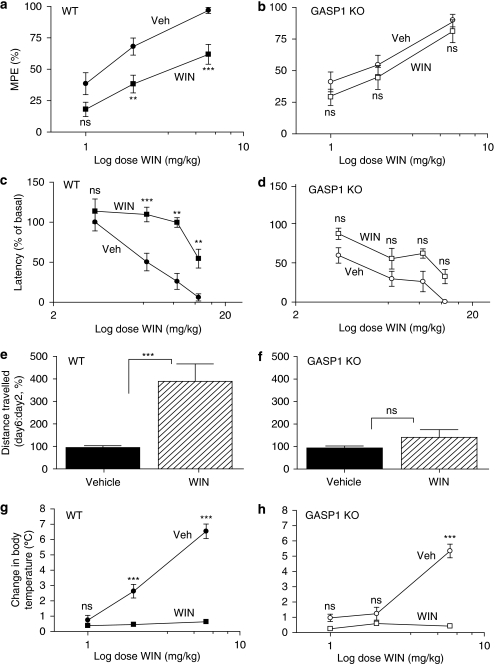
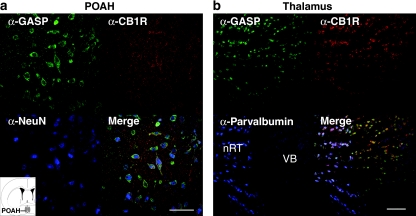
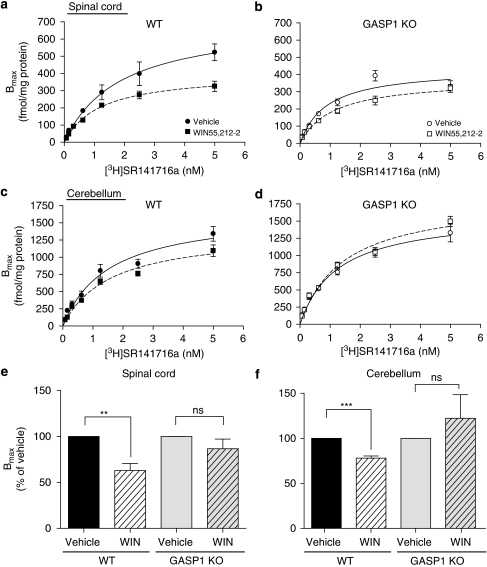
Cannabinoid agonists have shown some promise clinically as analgesics, in particular for cancer pain, in which they have the additional benefit of decreasing nausea. However, as for most other drugs, the long-term use of cannabinoids is limited by the development of tolerance. Several molecular mechanisms have been proposed to explain drug tolerance, including receptor downregulation. The cannabinoid 1 (CB1) receptors can be downregulated in vitro through an interaction with the G-protein-coupled receptor-associated sorting protein1, GASP1, that targets CB1 receptors for degradation after their agonist-mediated endocytosis. To investigate whether GASP1-mediated postendocytic sorting of the CB1 receptor contributes to tolerance to cannabinoid drugs in vivo, we generated a mouse with a disruption of GASP1. In wild-type mice, repeated administration of the cannabinoid agonist WIN55,212-2 promoted downregulation of CB1 receptor levels and concomitant tolerance to the effects of drug on antinociception, motor incoordination, and locomotor hypoactivity. In contrast, GASP1 knockout mice did not develop tolerance to any of these effects and showed no significant receptor downregulation. Taken together, this study provides evidence that GASP1 regulates CB1 receptor downregulation in vivo, and that postendocytic receptor trafficking has a key role in the development of tolerance to WIN55,212-2.
Figures
Similar articles
-
A molecular basis of analgesic tolerance to cannabinoids.J Neurosci. 2007 Apr 11;27(15):4165-77. doi: 10.1523/JNEUROSCI.5648-06.2007. J Neurosci. 2007. PMID: 17428994 Free PMC article.
-
PKCepsilon regulates behavioral sensitivity, binding and tolerance to the CB1 receptor agonist WIN55,212-2.Neuropsychopharmacology. 2009 Jun;34(7):1733-42. doi: 10.1038/npp.2008.230. Epub 2009 Jan 21. Neuropsychopharmacology. 2009. PMID: 19158669 Free PMC article.
-
Tolerance to cannabinoid-induced behaviors in mice treated chronically with ethanol.Psychopharmacology (Berl). 2012 Jan;219(1):137-47. doi: 10.1007/s00213-011-2387-0. Epub 2011 Jun 24. Psychopharmacology (Berl). 2012. PMID: 21701813 Free PMC article.
-
Regulation of cannabinoid CB1 receptors in the central nervous system by chronic cannabinoids.Crit Rev Neurobiol. 2003;15(2):91-119. doi: 10.1615/critrevneurobiol.v15.i2.10. Crit Rev Neurobiol. 2003. PMID: 14977366 Review.
-
Moving around the molecule: relationship between chemical structure and in vivo activity of synthetic cannabinoids.Life Sci. 2014 Feb 27;97(1):55-63. doi: 10.1016/j.lfs.2013.09.011. Epub 2013 Sep 23. Life Sci. 2014. PMID: 24071522 Free PMC article. Review.
Cited by
-
G protein-coupled receptor-associated sorting protein 1 regulates the postendocytic sorting of seven-transmembrane-spanning G protein-coupled receptors.Pharmacology. 2010;86(1):22-9. doi: 10.1159/000314161. Epub 2010 Jun 26. Pharmacology. 2010. PMID: 20693822 Free PMC article. Review.
-
SGIP1 is involved in regulation of emotionality, mood, and nociception and modulates in vivo signalling of cannabinoid CB1 receptors.Br J Pharmacol. 2021 Apr;178(7):1588-1604. doi: 10.1111/bph.15383. Epub 2021 Feb 27. Br J Pharmacol. 2021. PMID: 33491188 Free PMC article.
-
Cannabinoid receptor-interacting protein 1a modulates CB1 receptor signaling and regulation.Mol Pharmacol. 2015 Apr;87(4):747-65. doi: 10.1124/mol.114.096495. Epub 2015 Feb 5. Mol Pharmacol. 2015. PMID: 25657338 Free PMC article.
-
The expression of miR-21, HSP90a and gGASP-1 in serum of patients with lung cancer and their correlation with pathological subtypes.J Med Biochem. 2024 Jun 15;43(4):460-468. doi: 10.5937/jomb0-48051. J Med Biochem. 2024. PMID: 39139173 Free PMC article.
-
Cannabis use, abuse, and withdrawal: Cannabinergic mechanisms, clinical, and preclinical findings.J Neurochem. 2021 Jun;157(5):1674-1696. doi: 10.1111/jnc.15369. Epub 2021 May 16. J Neurochem. 2021. PMID: 33891706 Free PMC article. Review.
References
-
- Barna I, Till I, Haller J. Blood, adipose tissue and brain levels of the cannabinoid ligands WIN-55,212 and SR-141716A after their intraperitoneal injection in mice: compound-specific and area-specific distribution within the brain. Eur Neuropsychopharmacol. 2009;19:533–541. - PubMed
-
- Bass CE, Martin BR. Time course for the induction and maintenance of tolerance to Delta(9)-tetrahydrocannabinol in mice. Drug Alcohol Depend. 2000;60:113–119. - PubMed
-
- Boeuf J, Trigo JM, Moreau PH, Lecourtier L, Vogel E, Cassel JC, et al. Attenuated behavioural responses to acute and chronic cocaine in GASP-1-deficient mice. Eur J Neurosci. 2009;30:860–868. - PubMed
-
- Boulant JA. Hypothalamic mechanisms in thermoregulation. Fed Proc. 1981;40:2843–2850. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
