

TCR-peptide-MHC interactions in situ show accelerated kinetics and increased affinity
- PMID: 20164930
- PMCID: PMC3273423
- DOI: 10.1038/nature08746
TCR-peptide-MHC interactions in situ show accelerated kinetics and increased affinity
Abstract
The recognition of foreign antigens by T lymphocytes is essential to most adaptive immune responses. It is driven by specific T-cell antigen receptors (TCRs) binding to antigenic peptide-major histocompatibility complex (pMHC) molecules on other cells. If productive, these interactions promote the formation of an immunological synapse. Here we show that synaptic TCR-pMHC binding dynamics differ significantly from TCR-pMHC binding in solution. We used single-molecule microscopy and fluorescence resonance energy transfer (FRET) between fluorescently tagged TCRs and their cognate pMHC ligands to measure the kinetics of TCR-pMHC binding in situ. When compared with solution measurements, the dissociation of this complex was increased significantly (4-12-fold). Disruption of actin polymers reversed this effect, indicating that cytoskeletal dynamics destabilize this interaction directly or indirectly. Nevertheless, TCR affinity for pMHC was significantly elevated as the result of a large (about 100-fold) increase in the association rate, a likely consequence of complementary molecular orientation and clustering. In helper T cells, the CD4 molecule has been proposed to bind cooperatively with the TCR to the same pMHC complex. However, CD4 blockade had no effect on the synaptic TCR affinity, nor did it destabilize TCR-pMHC complexes, indicating that the TCR binds pMHC independently of CD4.
Figures
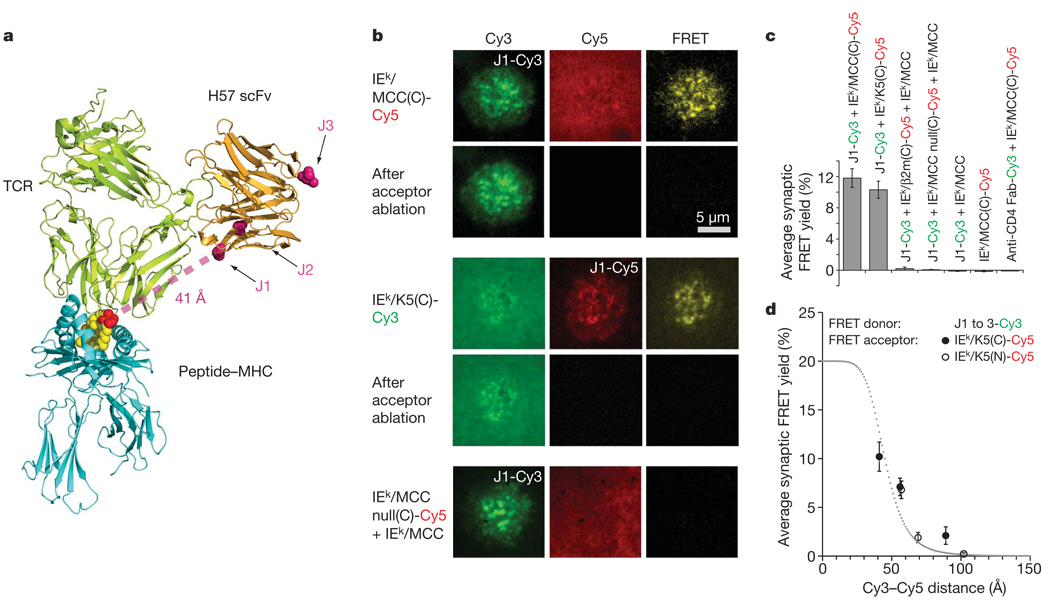
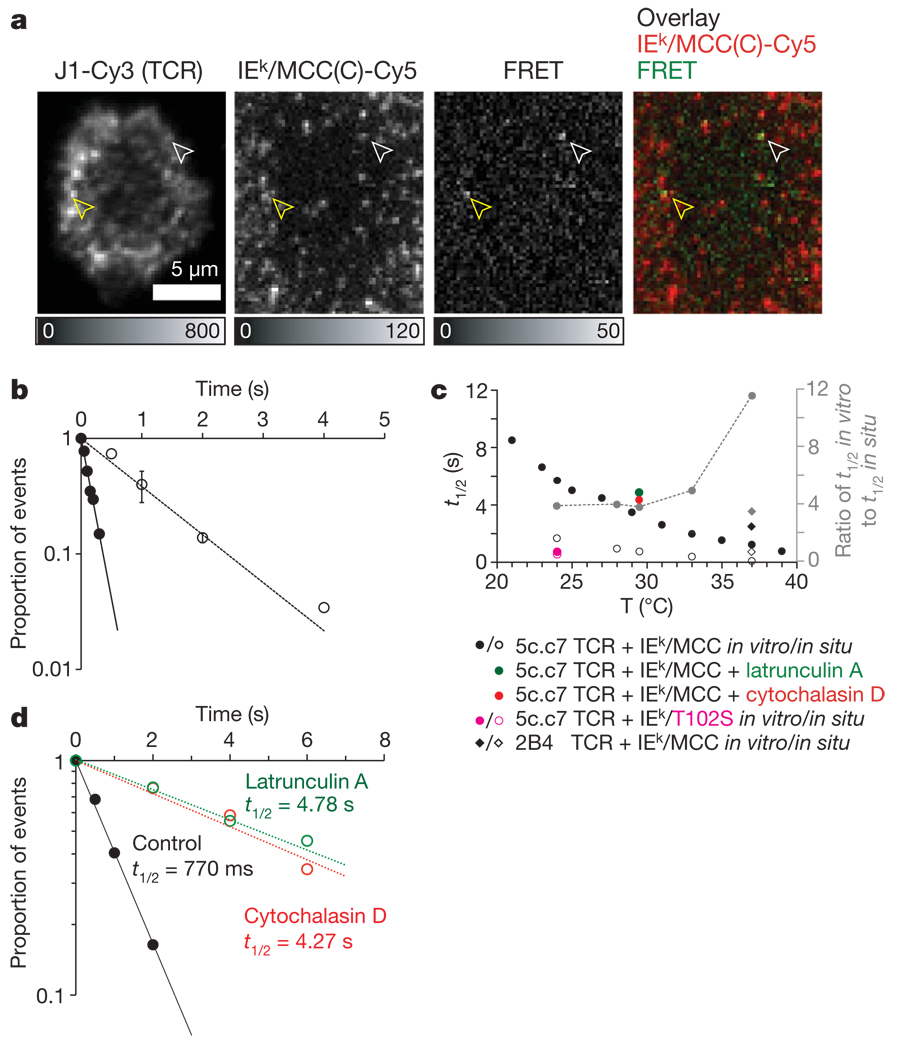
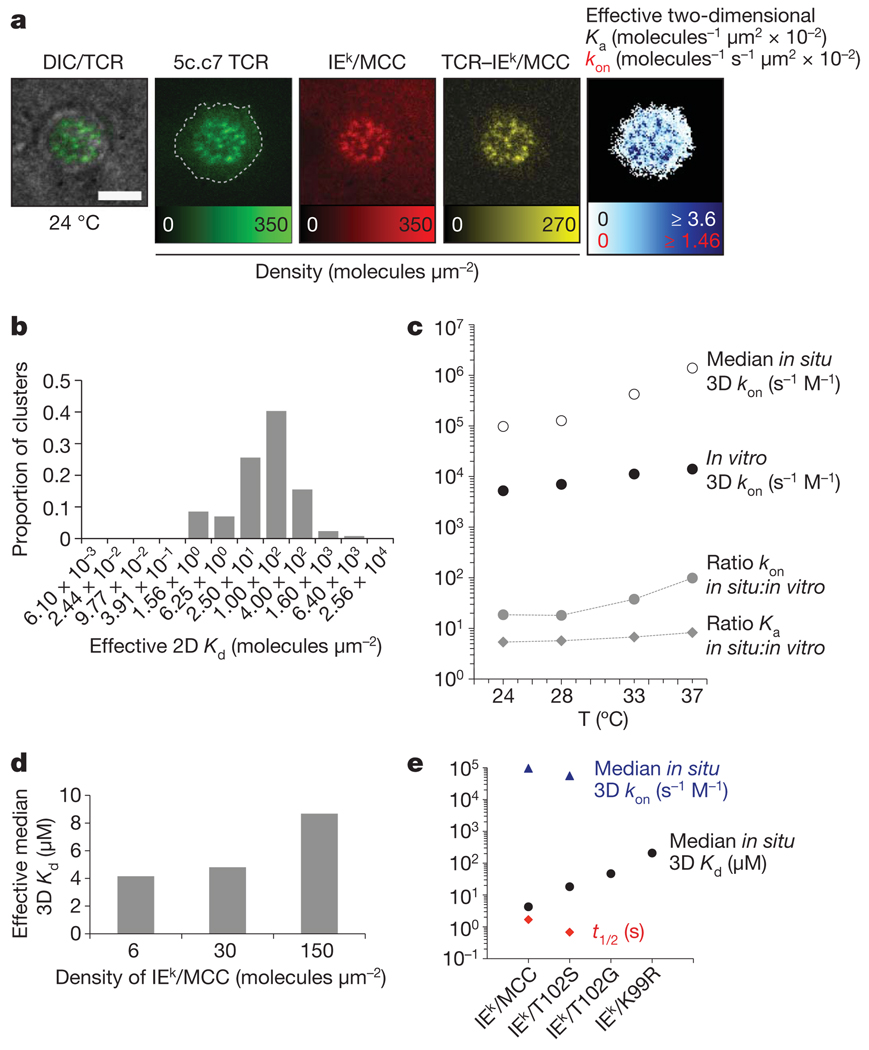
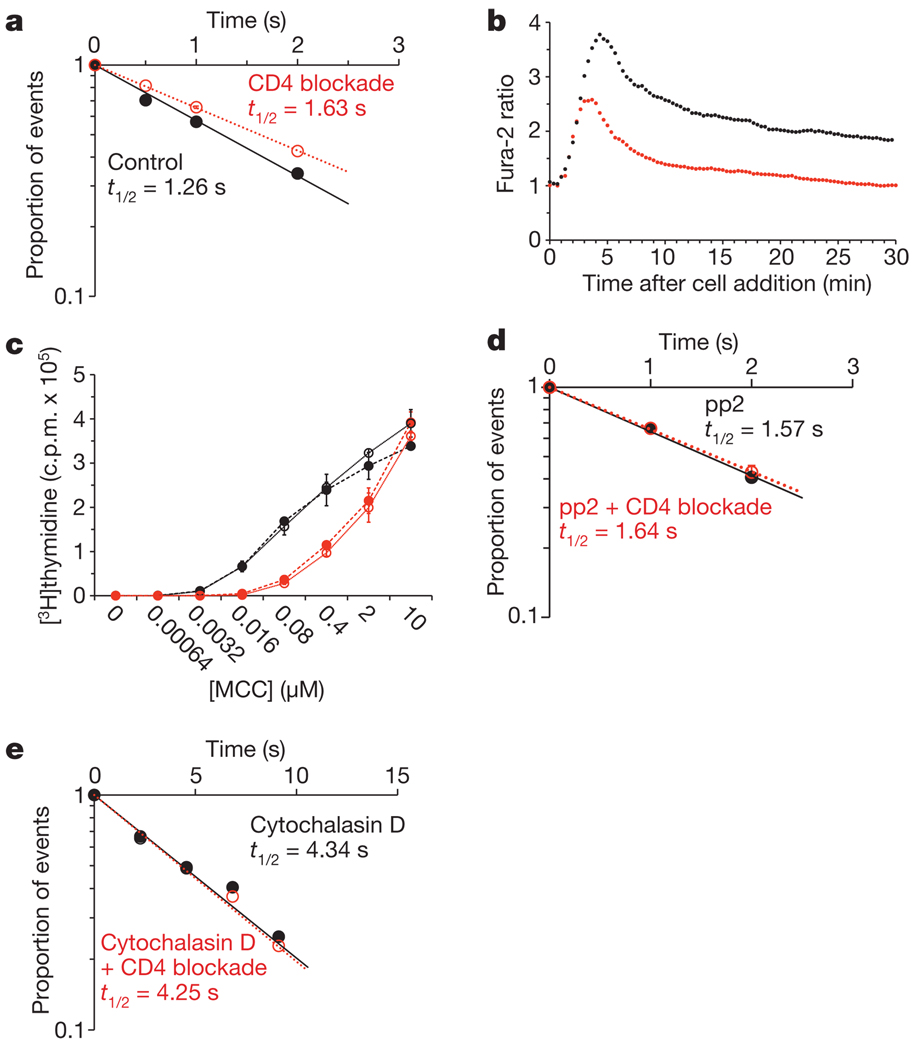
References
-
- Davis MM, et al. T cells as a self-referential, sensory organ. Annu. Rev. Immunol. 2007;25:681–695. - PubMed
-
- Grakoui A, et al. The immunological synapse: a molecular machine controlling cell activation. Science. 1999;285:221–227. - PubMed
-
- Monks CR, Freiberg BA, Kupfer H, Sciaky N, Kupfer A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature. 1998;395:82–86. - PubMed
-
- Corr M, et al. T cell receptor-MHC class I peptide interactions: affinity, kinetics, and specificity. Science. 1994;265:946–949. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
