

Insulin-like growth factor-binding protein-5-induced laminin gamma1 transcription requires filamin A
- PMID: 20167606
- PMCID: PMC2857142
- DOI: 10.1074/jbc.M109.061754
Insulin-like growth factor-binding protein-5-induced laminin gamma1 transcription requires filamin A
Abstract
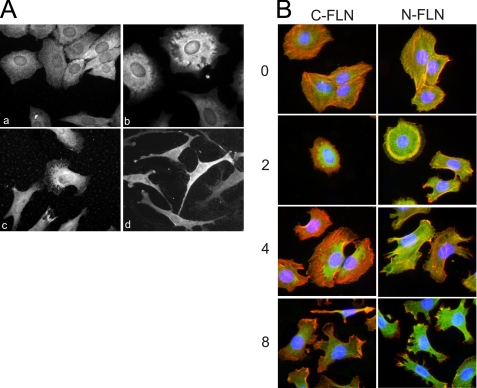
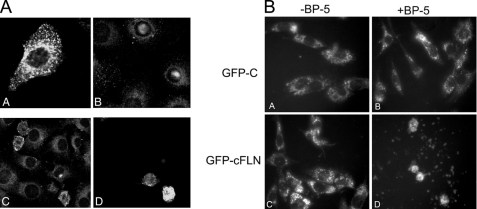
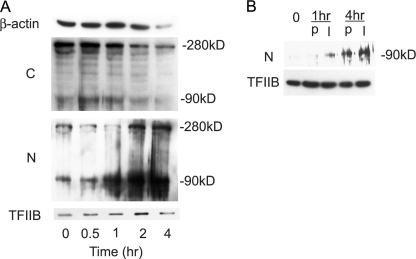
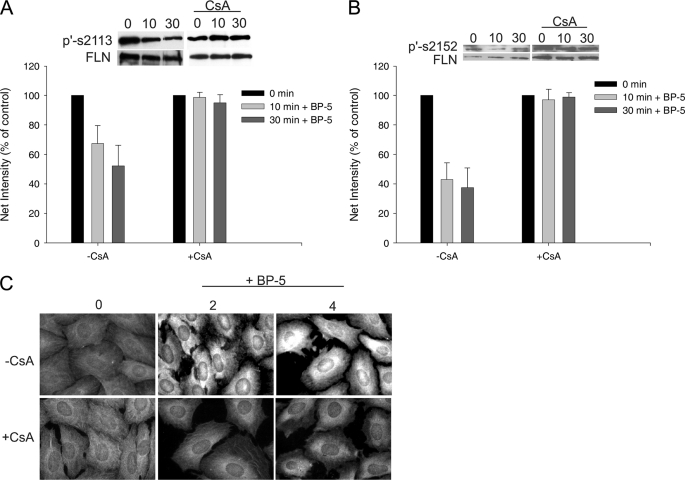
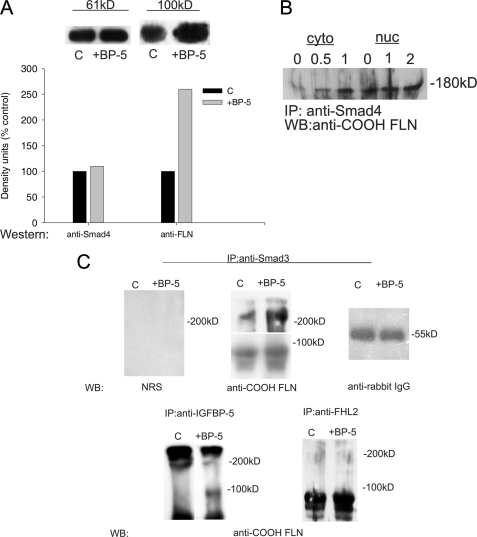
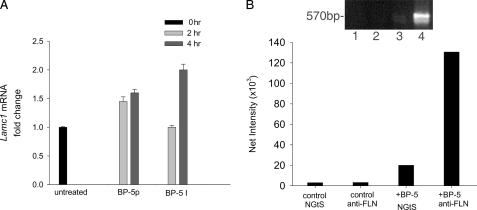
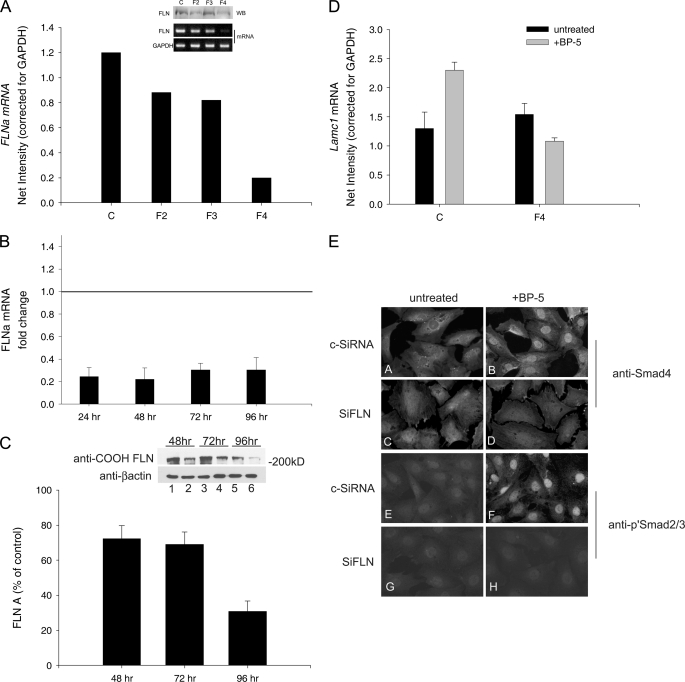
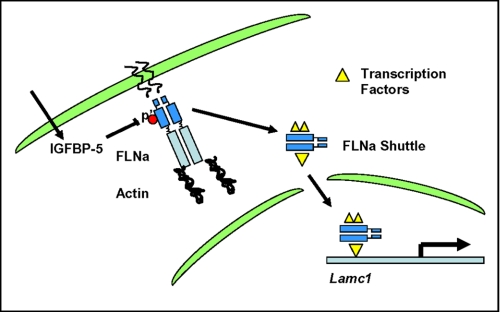
Insulin-like growth factor-binding protein-5 (IGFBP-5) has IGF-1-independent intranuclear effects that are poorly defined. Treatment of cells with IGFBP-5 induces migration, prevents apoptosis, and leads to increased laminin subunit transcription. Similarly, filamin A (FLNa), an actin-binding protein that participates in cell attachment, plays important additional roles in signal transduction and modulation of transcriptional responses. In this report, we show that IGFBP-5 leads to dephosphorylation of FLNa with subsequent FLNa cleavage. Following cleavage, there is enhanced recruitment of Smad3/4 to a C-terminal FLNa fragment with nuclear translocation and subsequent binding to the promoter region of the laminin gamma1 (lamc1) gene. FLNa knockdown prevents IGFBP-5-mediated increases in lamc1 transcription. These data indicate that IGFBP-5 induces formation of a FLNa-based nuclear shuttle that recruits transcription factors and regulates transcription of IGFBP-5 target genes. These studies provide new insights into the mechanisms whereby IGFBP-5 and FLNa exert intranuclear effects.
Figures
Similar articles
-
Nuclear versus cytoplasmic localization of filamin A in prostate cancer: immunohistochemical correlation with metastases.Clin Cancer Res. 2009 Feb 1;15(3):788-96. doi: 10.1158/1078-0432.CCR-08-1402. Clin Cancer Res. 2009. PMID: 19188148 Free PMC article.
-
Interaction of filamin A with the insulin receptor alters insulin-dependent activation of the mitogen-activated protein kinase pathway.J Biol Chem. 2003 Jul 18;278(29):27096-104. doi: 10.1074/jbc.M301003200. Epub 2003 May 6. J Biol Chem. 2003. PMID: 12734206
-
Ribosomal S6 kinase (RSK) regulates phosphorylation of filamin A on an important regulatory site.Mol Cell Biol. 2004 Apr;24(7):3025-35. doi: 10.1128/MCB.24.7.3025-3035.2004. Mol Cell Biol. 2004. PMID: 15024089 Free PMC article.
-
The filamins: organizers of cell structure and function.Cell Adh Migr. 2011 Mar-Apr;5(2):160-9. doi: 10.4161/cam.5.2.14401. Epub 2011 Mar 1. Cell Adh Migr. 2011. PMID: 21169733 Free PMC article. Review.
-
[Platelet filamin: a cytoskeletal protein involved in cell signal integration and function].Arch Cardiol Mex. 2006 Oct-Dec;76 Suppl 4:S67-75. Arch Cardiol Mex. 2006. PMID: 17469336 Review. Spanish.
Cited by
-
Regularization Methods for High-Dimensional Instrumental Variables Regression With an Application to Genetical Genomics.J Am Stat Assoc. 2015;110(509):270-288. doi: 10.1080/01621459.2014.908125. J Am Stat Assoc. 2015. PMID: 26392642 Free PMC article.
-
Hypoxia-induced and calpain-dependent cleavage of filamin A regulates the hypoxic response.Proc Natl Acad Sci U S A. 2014 Feb 18;111(7):2560-5. doi: 10.1073/pnas.1320815111. Epub 2014 Feb 3. Proc Natl Acad Sci U S A. 2014. PMID: 24550283 Free PMC article.
-
Development of self-healing hydrogels to support choroidal endothelial cell transplantation for the treatment of early age related macular degeneration.Acta Biomater. 2025 Mar 1;194:98-108. doi: 10.1016/j.actbio.2024.12.052. Epub 2024 Dec 20. Acta Biomater. 2025. PMID: 39710218
-
Molecular regulation of lifespan extension in fertile ant workers.Philos Trans R Soc Lond B Biol Sci. 2021 Apr 26;376(1823):20190736. doi: 10.1098/rstb.2019.0736. Epub 2021 Mar 8. Philos Trans R Soc Lond B Biol Sci. 2021. PMID: 33678017 Free PMC article.
-
The chromatin remodeling protein BRG1 regulates HSC-myofibroblast differentiation and liver fibrosis.Cell Death Dis. 2023 Dec 14;14(12):826. doi: 10.1038/s41419-023-06351-5. Cell Death Dis. 2023. PMID: 38092723 Free PMC article.
References
-
- Schneider M. R., Wolf E., Hoeflich A., Lahm H. (2002) J. Endocrinol. 172, 423–440 - PubMed
-
- Parker A., Clarke J. B., Busby W. H., Jr., Clemmons D. R. (1996) J. Biol. Chem. 271, 13523–13529 - PubMed
-
- Booth B. A., Boes M., Andress D. L., Dake B. L., Kiefer M. C., Maack C., Linhardt R. J., Bar K., Caldwell E. E., Weiler J., Bar R. S. (1995) Growth Regul. 5, 1–17 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
