

Cholesterol utilization in mycobacteria is controlled by two TetR-type transcriptional regulators: kstR and kstR2
- PMID: 20167624
- PMCID: PMC3068626
- DOI: 10.1099/mic.0.034538-0
Cholesterol utilization in mycobacteria is controlled by two TetR-type transcriptional regulators: kstR and kstR2
Abstract
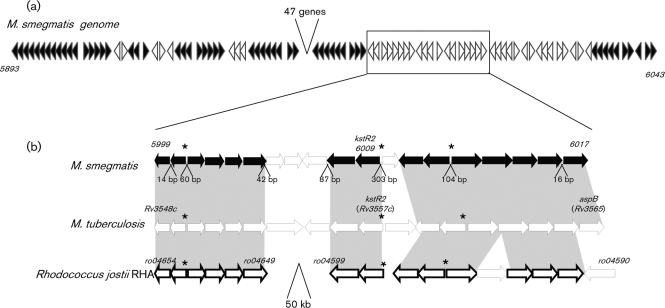
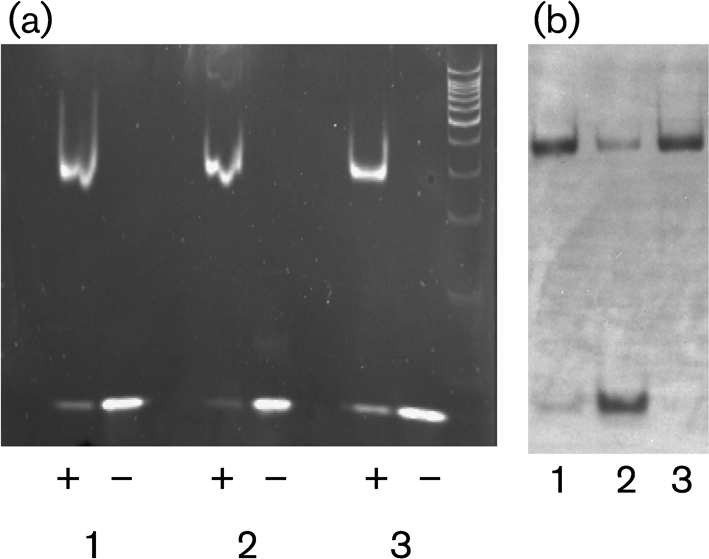
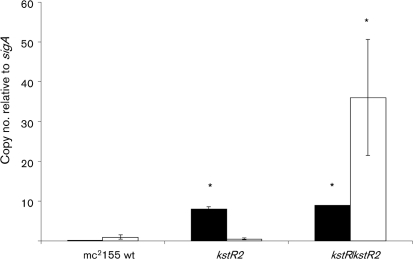
Mycobacterium tuberculosis is able to use a variety of carbon sources in vivo and current knowledge suggests that cholesterol is used as a carbon source during infection. The catabolized cholesterol is used both as an energy source (ATP generation) and as a source of precursor molecules for the synthesis of complex methyl-branched fatty acids. In previous studies, we described a TetR-type transcriptional repressor, kstR, that controls the expression of a number of genes involved in cholesterol catabolism. In this study, we describe a second TetR-type repressor, which we call kstR2. We knocked this gene out in Mycobacterium smegmatis and used microarrays and quantitative RT-PCR to examine the effects on gene expression. We identified a palindromic regulatory motif for KstR2, showed that this motif is present in three promoter regions in mycobacteria and rhodococcus, and demonstrated binding of purified KstR2 to the motif. Using a combination of motif location analysis, gene expression analysis and the examination of gene conservation, we suggest that kstR2 controls the expression of a 15 gene regulon. Like kstR, kstR2 and the kstR2 regulon are highly conserved among the actinomycetes and studies in rhodococcus suggest a role for these genes in cholesterol catabolism. The functional significance of the regulon and implications for the control of cholesterol utilization are discussed.
Figures

Similar articles
-
A highly conserved transcriptional repressor controls a large regulon involved in lipid degradation in Mycobacterium smegmatis and Mycobacterium tuberculosis.Mol Microbiol. 2007 Aug;65(3):684-99. doi: 10.1111/j.1365-2958.2007.05827.x. Mol Microbiol. 2007. PMID: 17635188 Free PMC article.
-
Characterization of the KstR2 regulator responsible of the lower cholesterol degradative pathway in Mycobacterium smegmatis.Environ Microbiol Rep. 2015 Feb;7(1):155-63. doi: 10.1111/1758-2229.12255. Epub 2015 Feb 12. Environ Microbiol Rep. 2015. PMID: 25511435
-
Regulation of the KstR2 regulon of Mycobacterium tuberculosis by a cholesterol catabolite.Mol Microbiol. 2013 Sep;89(6):1201-12. doi: 10.1111/mmi.12340. Epub 2013 Aug 14. Mol Microbiol. 2013. PMID: 23879670
-
Deciphering the transcriptional regulation of cholesterol catabolic pathway in mycobacteria: identification of the inducer of KstR repressor.J Biol Chem. 2014 Jun 20;289(25):17576-88. doi: 10.1074/jbc.M113.545715. Epub 2014 May 6. J Biol Chem. 2014. PMID: 24802756 Free PMC article.
-
GlnR-mediated regulation of KstR controls cholesterol catabolism in Mycobacterium smegmatis.Biotechnol Appl Biochem. 2022 Jun;69(3):1209-1216. doi: 10.1002/bab.2197. Epub 2021 May 27. Biotechnol Appl Biochem. 2022. PMID: 34008246
Cited by
-
Piperidinols that show anti-tubercular activity as inhibitors of arylamine N-acetyltransferase: an essential enzyme for mycobacterial survival inside macrophages.PLoS One. 2012;7(12):e52790. doi: 10.1371/journal.pone.0052790. Epub 2012 Dec 28. PLoS One. 2012. PMID: 23285185 Free PMC article.
-
Machine Learning of All Mycobacterium tuberculosis H37Rv RNA-seq Data Reveals a Structured Interplay between Metabolism, Stress Response, and Infection.mSphere. 2022 Apr 27;7(2):e0003322. doi: 10.1128/msphere.00033-22. Epub 2022 Mar 21. mSphere. 2022. PMID: 35306876 Free PMC article.
-
Host-Mycobacterium avium subsp. paratuberculosis interactome reveals a novel iron assimilation mechanism linked to nitric oxide stress during early infection.BMC Genomics. 2013 Oct 10;14:694. doi: 10.1186/1471-2164-14-694. BMC Genomics. 2013. PMID: 24112552 Free PMC article.
-
Purification, crystallization and preliminary X-ray crystallographic studies of KstR2 (ketosteroid regulatory protein) from Mycobacterium tuberculosis.Acta Crystallogr F Struct Biol Commun. 2014 Dec 1;70(Pt 12):1643-5. doi: 10.1107/S2053230X14023589. Epub 2014 Nov 14. Acta Crystallogr F Struct Biol Commun. 2014. PMID: 25484217 Free PMC article.
-
Delineation of Steroid-Degrading Microorganisms through Comparative Genomic Analysis.mBio. 2016 Mar 8;7(2):e00166. doi: 10.1128/mBio.00166-16. mBio. 2016. PMID: 26956583 Free PMC article.
References
-
- Bailey, T. L. & Elkan, C. (1994). Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol 2, 28–36. - PubMed
-
- Bailey, T. L. & Gribskov, M. (1998). Combining evidence using p-values: application to sequence homology searches. Bioinformatics 14, 48–54. - PubMed
-
- Benjamini, Y. (1995). Controlling false discovery rate: a practical and powerful approach to multiple testing. J Roy Stat Soc B 57, 289–300.
-
- Brzostek, A., Dziadek, B., Rumijowska-Galewicz, A., Pawelczyk, J. & Dziadek, J. (2007). Cholesterol oxidase is required for virulence of Mycobacterium tuberculosis. FEMS Microbiol Lett 275, 106–112. - PubMed
-
- Camus, J. C., Pryor, M. J., Medigue, C. & Cole, S. T. (2002). Re-annotation of the genome sequence of Mycobacterium tuberculosis H37Rv. Microbiology 148, 2967–2973. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous