

The NoRC complex mediates the heterochromatin formation and stability of silent rRNA genes and centromeric repeats
- PMID: 20168299
- PMCID: PMC2905252
- DOI: 10.1038/emboj.2010.17
The NoRC complex mediates the heterochromatin formation and stability of silent rRNA genes and centromeric repeats
Erratum in
- EMBO J. 2010 Jul 7;29(13):2253
Abstract
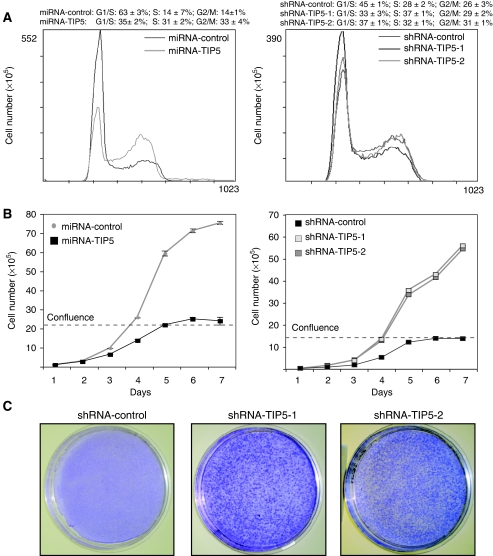
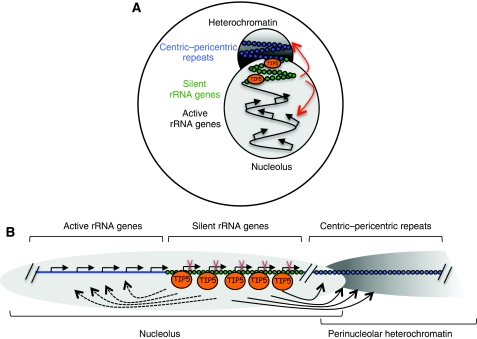
Maintenance of specific heterochromatic domains is crucial for genome stability. In eukaryotic cells, a fraction of the tandem-repeated ribosomal RNA (rRNA) genes is organized in the heterochromatic structures. The principal determinant of rDNA silencing is the nucleolar remodelling complex, NoRC, that consists of TIP5 (TTF-1-interacting protein-5) and the ATPase SNF2h. Here we showed that TIP5 not only mediates the establishment of rDNA silencing but also the formation of perinucleolar heterochromatin that contains centric and pericentric repeats. Our data indicated that the TIP5-mediated heterochromatin is indispensable for stability of silent rRNA genes and of major and minor satellite repeats. Moreover, depletion of TIP5 impairs rDNA silencing, upregulates rDNA transcription levels and induces cell transformation. These findings point to a role of TIP5 in protecting genome stability and suggest that it can play a role in the cellular transformation process.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
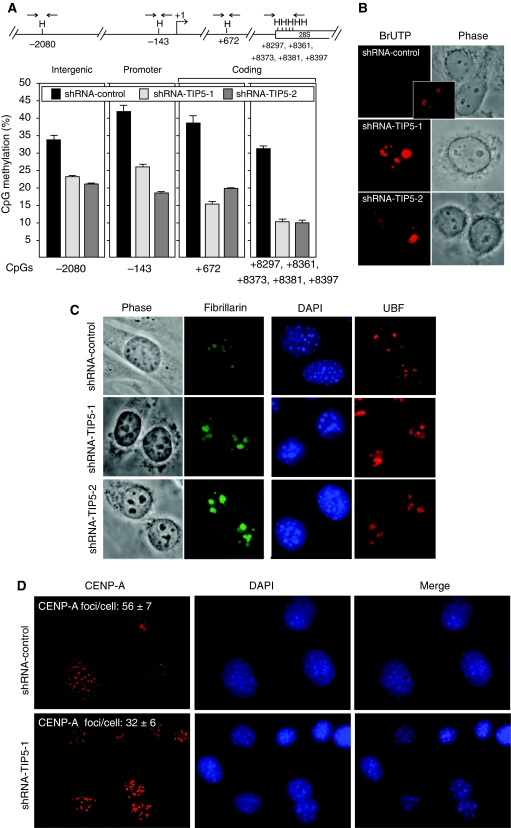
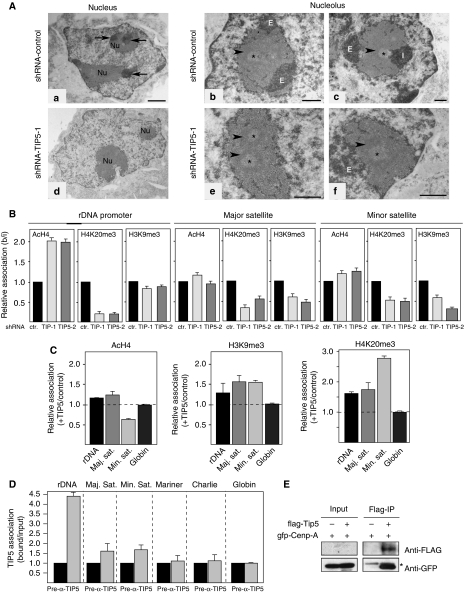
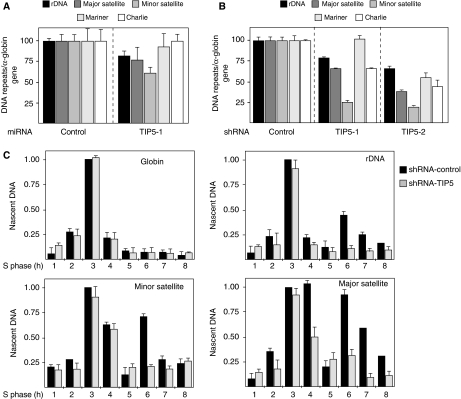
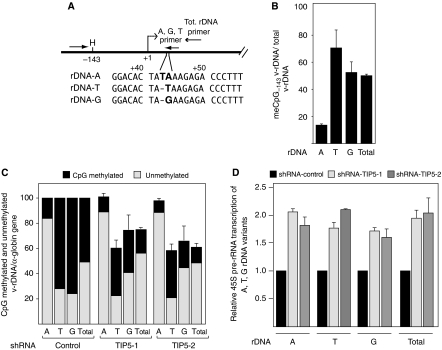
References
-
- Akhmanova A, Verkerk T, Langeveld A, Grosveld F, Galiart N (2000) Characterisation of transcriptionally active and inactive chromatin domains in neurons. J Cell Sci 24: 4463–4474 - PubMed
-
- Arnheim N, Southern EM (1977) Heterogeneity of the ribosomal genes in mice and men. Cell 11: 363–370 - PubMed
-
- Black BE, Bassett EA (2008) The histone variant CENP-A and centromere specification. Curr Opin Cell Biol 20: 91–100 - PubMed
-
- Boisvert FM, van Koningsbruggen S, Navascués J, Lamond AI (2007) The multifunctional nucleolus. Nat Rev Mol Cell Biol 8: 574–585 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
