

Vacuolar ATPase regulates surfactant secretion in rat alveolar type II cells by modulating lamellar body calcium
- PMID: 20169059
- PMCID: PMC2821907
- DOI: 10.1371/journal.pone.0009228
Vacuolar ATPase regulates surfactant secretion in rat alveolar type II cells by modulating lamellar body calcium
Abstract
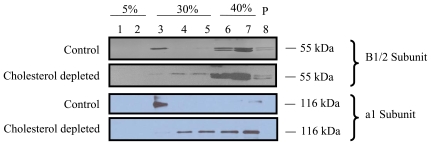
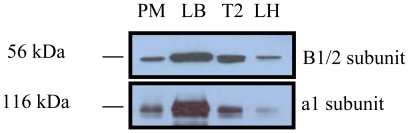
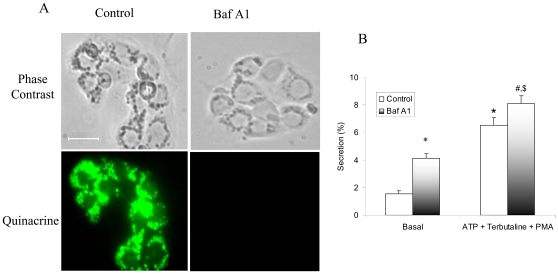
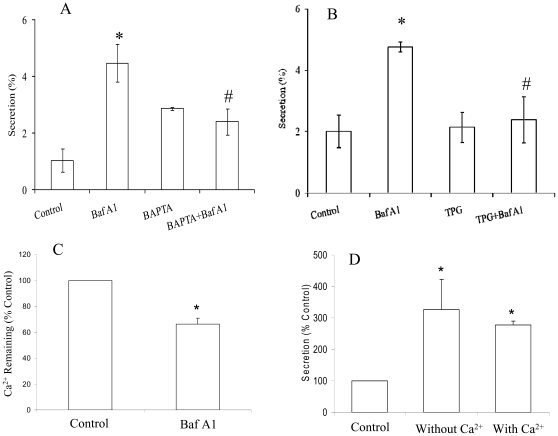
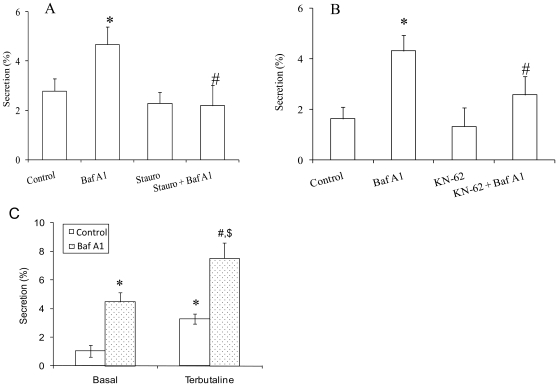
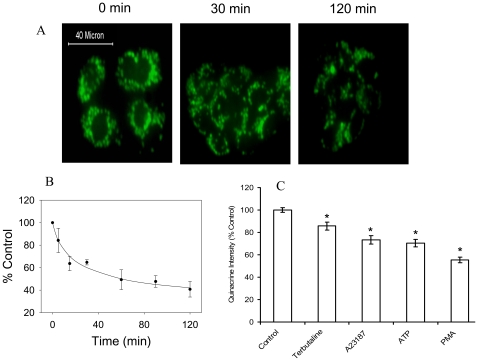
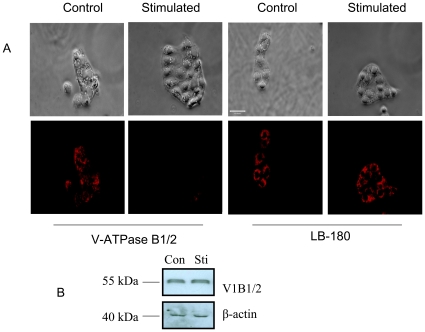
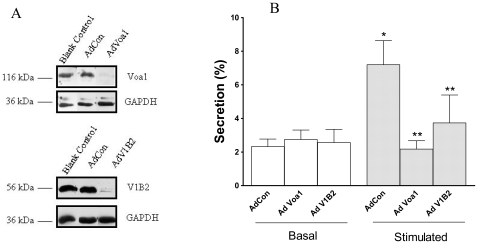
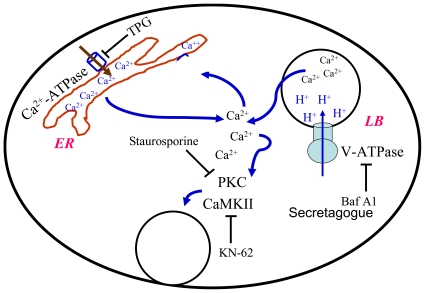
Lung surfactant reduces surface tension and maintains the stability of alveoli. How surfactant is released from alveolar epithelial type II cells is not fully understood. Vacuolar ATPase (V-ATPase) is the enzyme responsible for pumping H(+) into lamellar bodies and is required for the processing of surfactant proteins and the packaging of surfactant lipids. However, its role in lung surfactant secretion is unknown. Proteomic analysis revealed that vacuolar ATPase (V-ATPase) dominated the alveolar type II cell lipid raft proteome. Western blotting confirmed the association of V-ATPase a1 and B1/2 subunits with lipid rafts and their enrichment in lamellar bodies. The dissipation of lamellar body pH gradient by Bafilomycin A1 (Baf A1), an inhibitor of V-ATPase, increased surfactant secretion. Baf A1-stimulated secretion was blocked by the intracellular Ca(2+) chelator, BAPTA-AM, the protein kinase C (PKC) inhibitor, staurosporine, and the Ca(2+)/calmodulin-dependent protein kinase II (CaMKII), KN-62. Baf A1 induced Ca(2+) release from isolated lamellar bodies. Thapsigargin reduced the Baf A1-induced secretion, indicating cross-talk between lamellar body and endoplasmic reticulum Ca(2+) pools. Stimulation of type II cells with surfactant secretagogues dissipated the pH gradient across lamellar bodies and disassembled the V-ATPase complex, indicating the physiological relevance of the V-ATPase-mediated surfactant secretion. Finally, silencing of V-ATPase a1 and B2 subunits decreased stimulated surfactant secretion, indicating that these subunits were crucial for surfactant secretion. We conclude that V-ATPase regulates surfactant secretion via an increased Ca(2+) mobilization from lamellar bodies and endoplasmic reticulum, and the activation of PKC and CaMKII. Our finding revealed a previously unrealized role of V-ATPase in surfactant secretion.
Conflict of interest statement
Figures

Similar articles
-
Secretagogues of lung surfactant increase annexin A7 localization with ABCA3 in alveolar type II cells.Biochim Biophys Acta. 2011 Dec;1813(12):2017-25. doi: 10.1016/j.bbamcr.2011.07.022. Epub 2011 Sep 2. Biochim Biophys Acta. 2011. PMID: 21911013 Free PMC article.
-
H+-and K+-dependence of Ca2+ uptake in lung lamellar bodies.J Membr Biol. 2000 Mar 1;174(1):41-51. doi: 10.1007/s002320001030. J Membr Biol. 2000. PMID: 10741431
-
Binding of annexins to lung lamellar bodies and the PMA-stimulated secretion of annexin V from alveolar type II cells.J Biochem. 2001 Sep;130(3):449-55. doi: 10.1093/oxfordjournals.jbchem.a003005. J Biochem. 2001. PMID: 11530022
-
Regulation of surfactant secretion in alveolar type II cells.Am J Physiol Lung Cell Mol Physiol. 2007 Aug;293(2):L259-71. doi: 10.1152/ajplung.00112.2007. Epub 2007 May 11. Am J Physiol Lung Cell Mol Physiol. 2007. PMID: 17496061 Review.
-
Aspects of metabolism and storage of pulmonary surfactant: experiments with isolated type II pneumocytes and lamellar bodies.Eur J Respir Dis Suppl. 1987;153:182-8. Eur J Respir Dis Suppl. 1987. PMID: 3322861 Review.
Cited by
-
Long-chain acyl-CoA dehydrogenase deficiency as a cause of pulmonary surfactant dysfunction.J Biol Chem. 2014 Apr 11;289(15):10668-10679. doi: 10.1074/jbc.M113.540260. Epub 2014 Mar 3. J Biol Chem. 2014. PMID: 24591516 Free PMC article.
-
Oxidized Ferric and Ferryl Forms of Hemoglobin Trigger Mitochondrial Dysfunction and Injury in Alveolar Type I Cells.Am J Respir Cell Mol Biol. 2016 Aug;55(2):288-98. doi: 10.1165/rcmb.2015-0197OC. Am J Respir Cell Mol Biol. 2016. PMID: 26974230 Free PMC article.
-
Regulation of luminal acidification by the V-ATPase.Physiology (Bethesda). 2013 Sep;28(5):318-29. doi: 10.1152/physiol.00007.2013. Physiology (Bethesda). 2013. PMID: 23997191 Free PMC article. Review.
-
Acidic Enough for a Healthy Breath.Am J Respir Cell Mol Biol. 2024 Oct;71(4):383-385. doi: 10.1165/rcmb.2024-0237ED. Am J Respir Cell Mol Biol. 2024. PMID: 38889348 Free PMC article. No abstract available.
-
Adhesion-GPCR Gpr116 (ADGRF5) expression inhibits renal acid secretion.Proc Natl Acad Sci U S A. 2020 Oct 20;117(42):26470-26481. doi: 10.1073/pnas.2007620117. Epub 2020 Oct 1. Proc Natl Acad Sci U S A. 2020. PMID: 33004624 Free PMC article.
References
-
- Pelkmans L, Helenius A. Endocytosis via caveolae. Traffic. 2002;3:311–320. - PubMed
-
- Allen JA, Halverson-Tamboli RA, Rasenick MM. Lipid raft microdomains and neurotransmitter signalling. Nat Rev Neurosci. 2007;8:128–140. - PubMed
-
- Schuck S, Simons K. Polarized sorting in epithelial cells: raft clustering and the biogenesis of the apical membrane. J Cell Sci. 2004;117:5955–5964. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
