

Pathogen entrapment by transglutaminase--a conserved early innate immune mechanism
- PMID: 20169185
- PMCID: PMC2820530
- DOI: 10.1371/journal.ppat.1000763
Pathogen entrapment by transglutaminase--a conserved early innate immune mechanism
Abstract
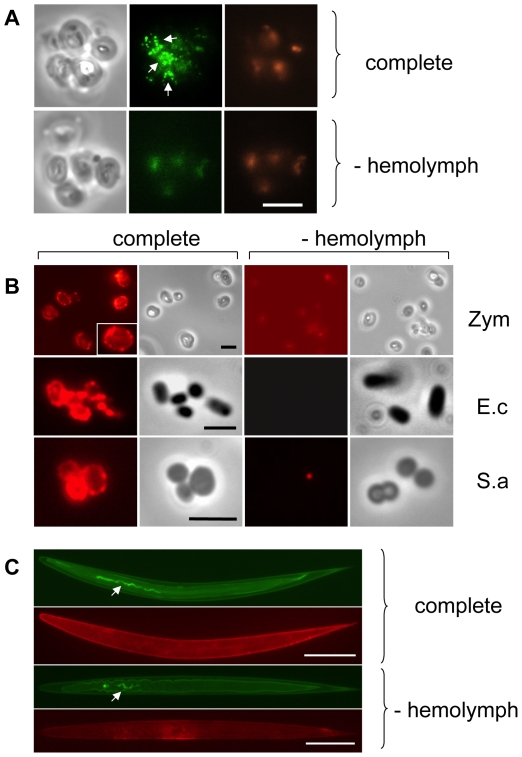
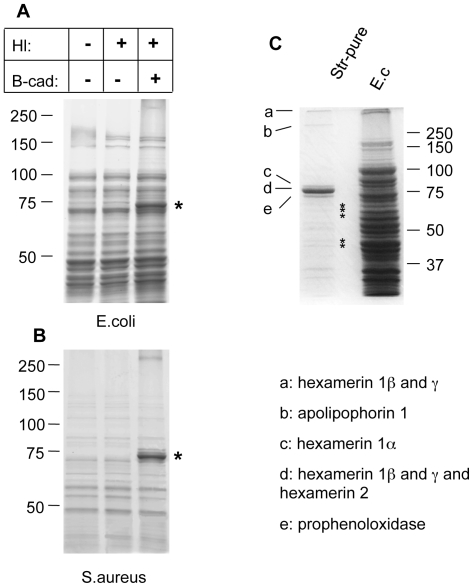
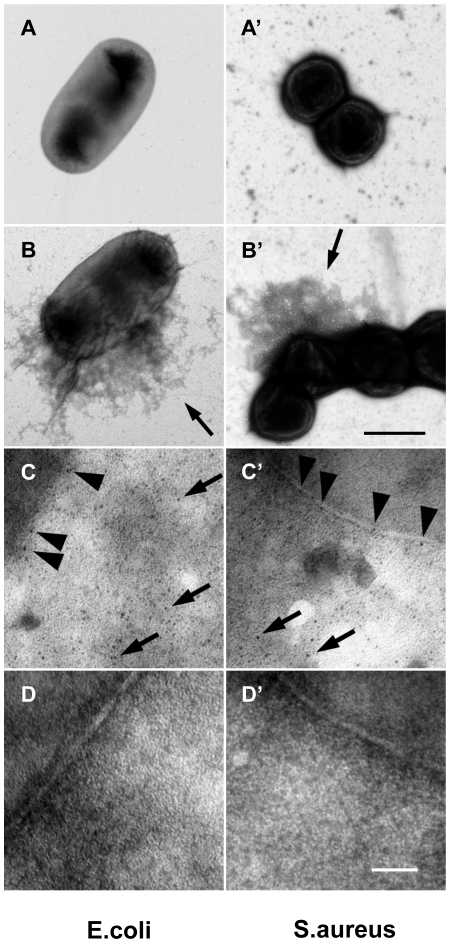
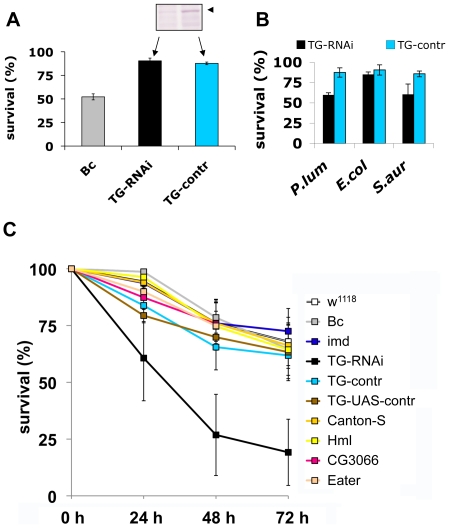
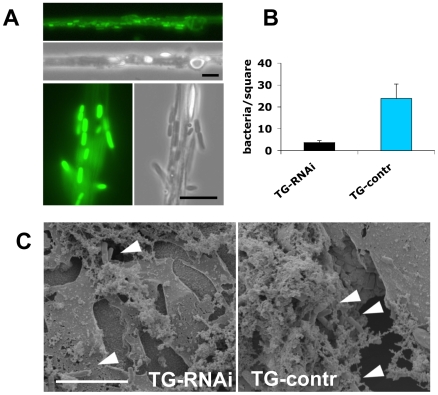
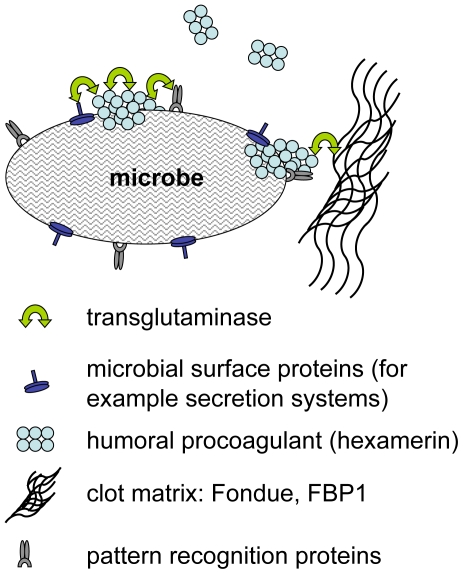
Clotting systems are required in almost all animals to prevent loss of body fluids after injury. Here, we show that despite the risks associated with its systemic activation, clotting is a hitherto little appreciated branch of the immune system. We compared clotting of human blood and insect hemolymph to study the best-conserved component of clotting systems, namely the Drosophila enzyme transglutaminase and its vertebrate homologue Factor XIIIa. Using labelled artificial substrates we observe that transglutaminase activity from both Drosophila hemolymph and human blood accumulates on microbial surfaces, leading to their sequestration into the clot. Using both a human and a natural insect pathogen we provide functional proof for an immune function for transglutaminase (TG). Drosophila larvae with reduced TG levels show increased mortality after septic injury. The same larvae are also more susceptible to a natural infection involving entomopathogenic nematodes and their symbiotic bacteria while neither phagocytosis, phenoloxidase or-as previously shown-the Toll or imd pathway contribute to immunity. These results firmly establish the hemolymph/blood clot as an important effector of early innate immunity, which helps to prevent septic infections. These findings will help to guide further strategies to reduce the damaging effects of clotting and enhance its beneficial contribution to immune reactions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Sun H. The interaction between pathogens and the host coagulation system. Physiology (Bethesda) 2006;21:281–288. - PubMed
-
- Rowley AF, Ratcliffe NA. The granular cells of Galleria mellonella during clotting and phagocytic reactions in vitro. Tissue and Cell. 1976;8:437–446. - PubMed
-
- Bidla G, Lindgren M, Theopold U, Dushay MS. Hemolymph coagulation and phenoloxidase in Drosophila larvae. Dev Comp Immunol. 2005;29:669–679. - PubMed
-
- Scherfer C, Qazi MR, Takahashi K, Ueda R, Dushay MS, et al. The Toll immune-regulated Drosophila protein Fondue is involved in hemolymph clotting and puparium formation. Dev Biol. 2006;295:156–163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
