

The early-onset torsion dystonia-associated protein, torsinA, displays molecular chaperone activity in vitro
- PMID: 20169475
- PMCID: PMC3006618
- DOI: 10.1007/s12192-010-0173-2
The early-onset torsion dystonia-associated protein, torsinA, displays molecular chaperone activity in vitro
Abstract
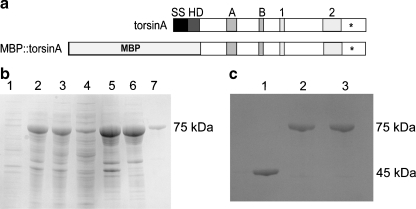
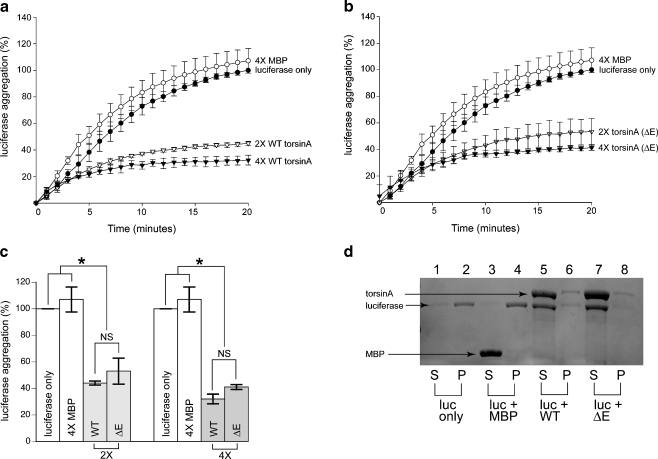
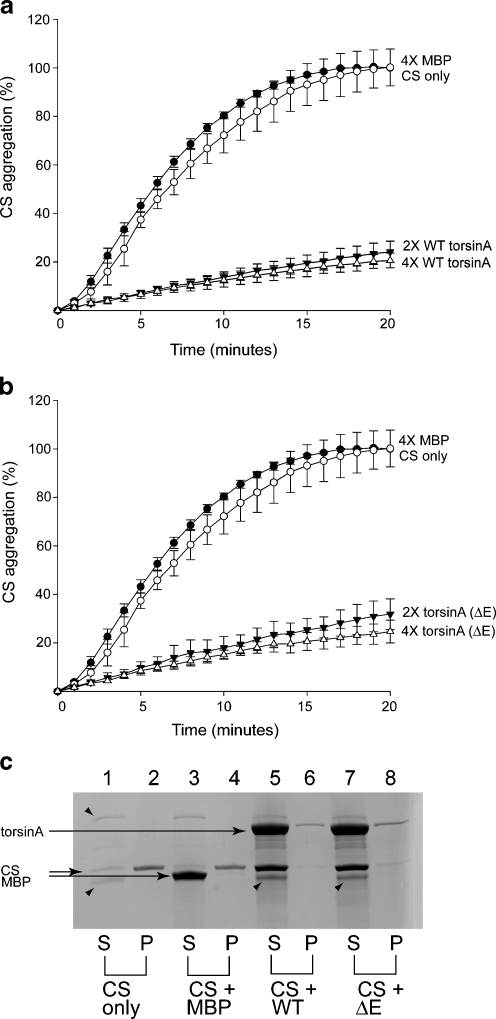
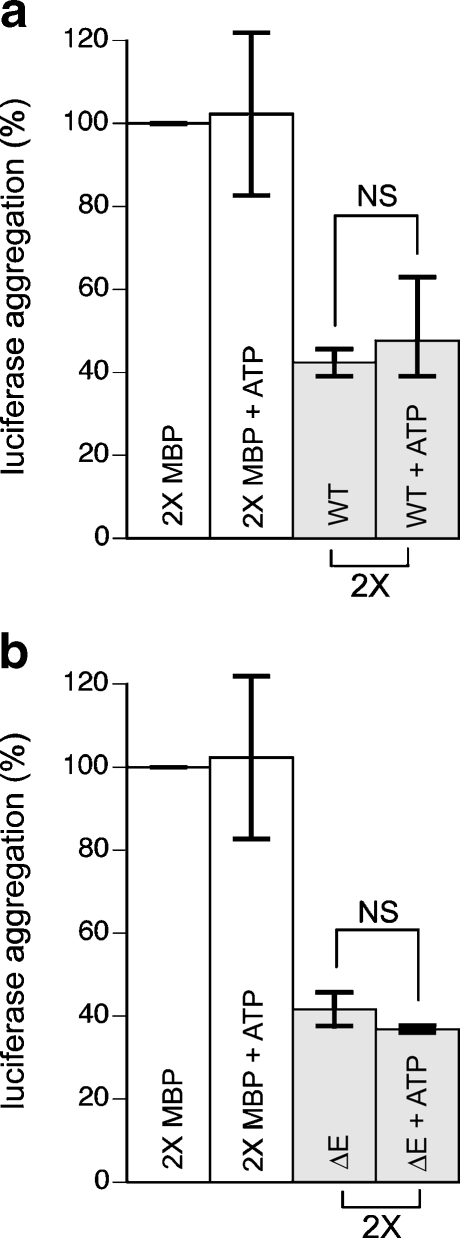
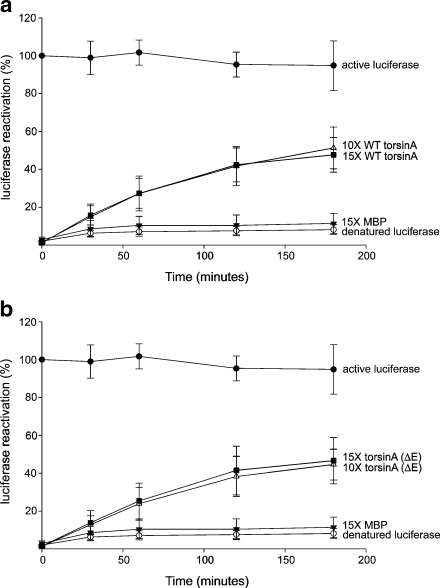
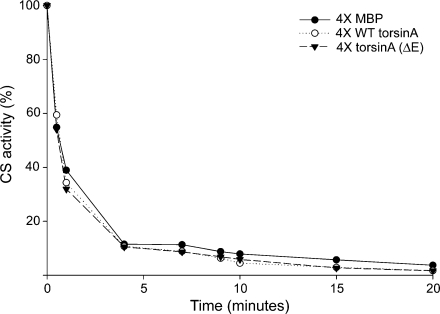
TorsinA is a member of the AAA+ ATPase family of proteins and, notably, is the only known ATPase localized to the ER lumen. It has been suggested to act as a molecular chaperone, while a mutant form associated with early-onset torsion dystonia, a dominantly inherited movement disorder, appears to result in a net loss of function in vivo. Thus far, no studies have examined the chaperone activity of torsinA in vitro. Here we expressed and purified both wild-type (WT) and mutant torsinA fusion proteins in bacteria and examined their ability to function as molecular chaperones by monitoring suppression of luciferase and citrate synthase (CS) aggregation. We also assessed their ability to hold proteins in an intermediate state for refolding. As measured by light scattering and SDS-PAGE, both WT and mutant torsinA effectively, and similarly, suppressed protein aggregation compared to controls. This function was not further enhanced by the presence of ATP. Further, we found that while neither form of torsinA could protect CS from heat-induced inactivation, they were both able to reactivate luciferase when ATP and rabbit reticulocyte lysate were added. This suggests that torsinA holds luciferase in an intermediate state, which can then be refolded in the presence of other chaperones. These data provide conclusive evidence that torsinA acts as a molecular chaperone in vitro and suggests that early-onset torsion dystonia is likely not a consequence of a loss in torsinA chaperone activity but might be an outcome of insufficient torsinA localization at the ER to manage protein folding or trafficking.
Figures
References
-
- Alexandrov A, Dutta K, Pascal SM. MBP fusion protein with a viral protease cleavage site: one-step cleavage/purification of insoluble proteins. Biotechniques. 2001;30(6):1194–1198. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
