

A central role for the WH2 domain of Srv2/CAP in recharging actin monomers to drive actin turnover in vitro and in vivo
- PMID: 20169536
- PMCID: PMC2857556
- DOI: 10.1002/cm.20429
A central role for the WH2 domain of Srv2/CAP in recharging actin monomers to drive actin turnover in vitro and in vivo
Abstract
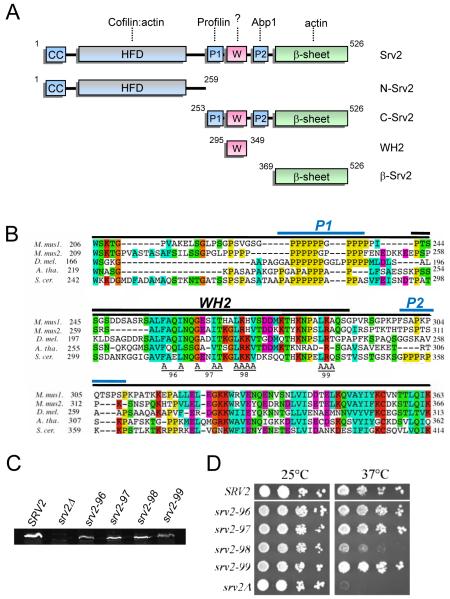
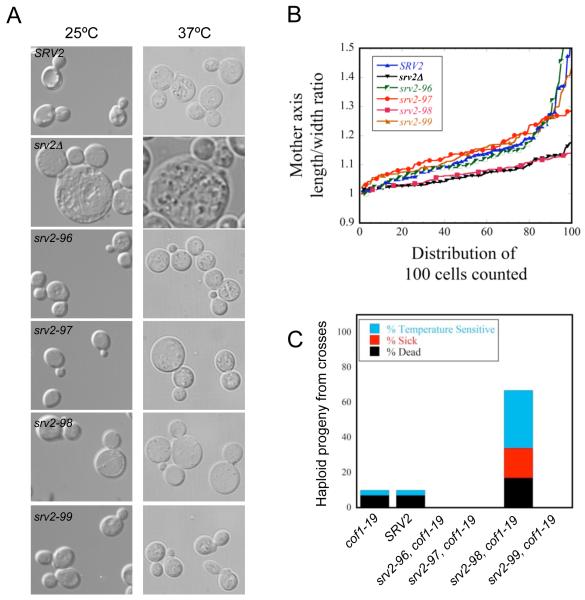
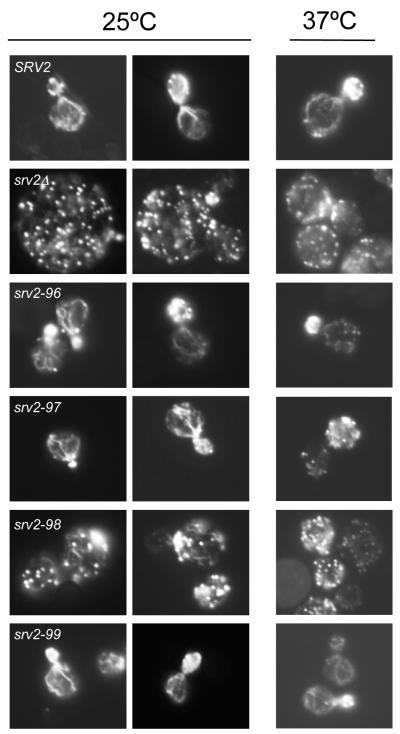
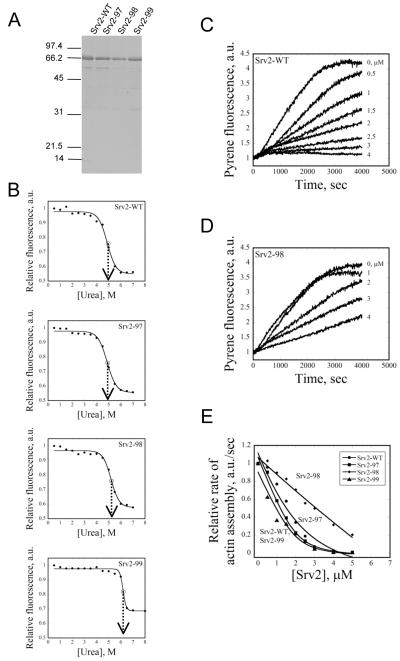
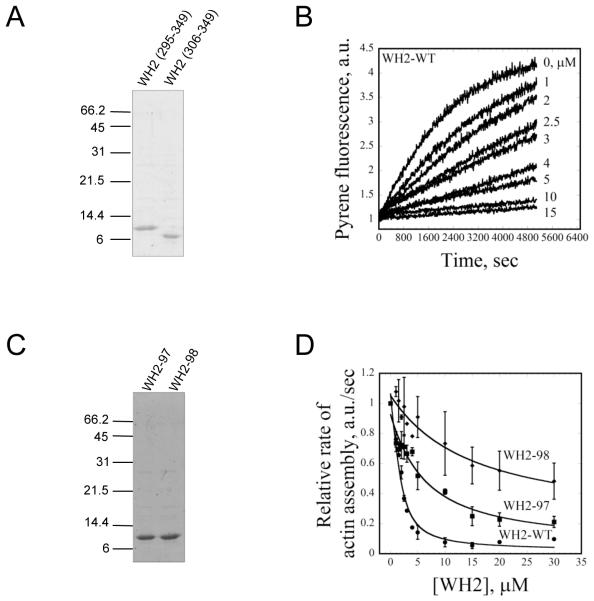
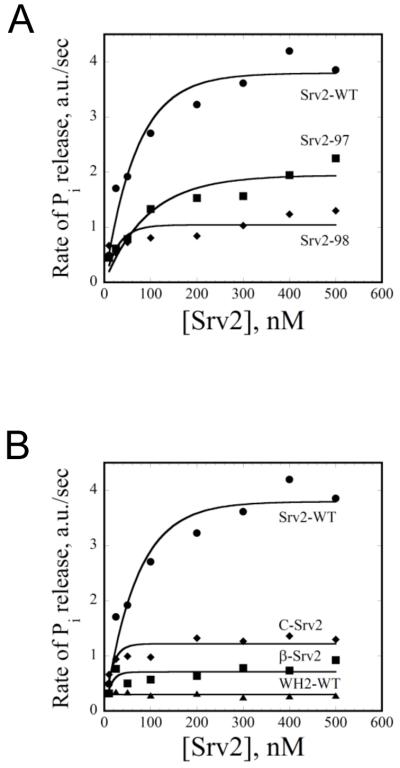
Cellular processes propelled by actin polymerization require rapid disassembly of filaments, and then efficient recycling of ADF/cofilin-bound ADP-actin monomers back to an assembly-competent ATP-bound state. How monomer recharging is regulated in vivo is still not well understood, but recent work suggests the involvement of the ubiquitous actin-monomer binding protein Srv2/CAP. To better understand Srv2/CAP mechanism, we explored the contribution of its WH2 domain, the function of which has remained highly elusive. We found that the WH2 domain binds to actin monomers and, unlike most other WH2 domains, exhibits similar binding affinity for ATP-actin and ADP-actin (K(d) approximately 1.5 microM). Mutations in the WH2 domain that impair actin binding disrupt the ability of purified full-length Srv2/CAP to catalyze nucleotide exchange on ADF/cofilin-bound actin monomers and accelerate actin turnover in vitro. The same mutations impair Srv2/CAP function in vivo in regulating actin organization, cell growth, and cell morphogenesis. Thus, normal cell growth and organization depend on the ability of Srv2/CAP to recharge actin monomers, and the WH2 domain plays a central role in this process. Our data also reveal that while most isolated WH2 domains inhibit nucleotide exchange on actin, WH2 domains in the context of intact proteins can help promote nucleotide exchange.
2010 Wiley-Liss, Inc.
Figures


Similar articles
-
Reconstitution and dissection of the 600-kDa Srv2/CAP complex: roles for oligomerization and cofilin-actin binding in driving actin turnover.J Biol Chem. 2009 Apr 17;284(16):10923-34. doi: 10.1074/jbc.M808760200. Epub 2009 Feb 6. J Biol Chem. 2009. PMID: 19201756 Free PMC article.
-
Coordinated regulation of actin filament turnover by a high-molecular-weight Srv2/CAP complex, cofilin, profilin, and Aip1.Curr Biol. 2003 Dec 16;13(24):2159-69. doi: 10.1016/j.cub.2003.11.051. Curr Biol. 2003. PMID: 14680631
-
Mechanism and biological role of profilin-Srv2/CAP interaction.J Cell Sci. 2007 Apr 1;120(Pt 7):1225-34. doi: 10.1242/jcs.000158. J Cell Sci. 2007. PMID: 17376963
-
WH2 domain: a small, versatile adapter for actin monomers.FEBS Lett. 2002 Feb 20;513(1):92-7. doi: 10.1016/s0014-5793(01)03242-2. FEBS Lett. 2002. PMID: 11911886 Review.
-
The role of cyclase-associated protein in regulating actin filament dynamics - more than a monomer-sequestration factor.J Cell Sci. 2013 Aug 1;126(Pt 15):3249-58. doi: 10.1242/jcs.128231. J Cell Sci. 2013. PMID: 23908377 Free PMC article. Review.
Cited by
-
Profilin connects actin assembly with microtubule dynamics.Mol Biol Cell. 2016 Aug 1;27(15):2381-93. doi: 10.1091/mbc.E15-11-0799. Epub 2016 Jun 15. Mol Biol Cell. 2016. PMID: 27307590 Free PMC article.
-
Multicomponent depolymerization of actin filament pointed ends by cofilin and cyclase-associated protein depends upon filament age.bioRxiv [Preprint]. 2024 Apr 15:2024.04.15.589566. doi: 10.1101/2024.04.15.589566. bioRxiv. 2024. Update in: Eur J Cell Biol. 2024 Jun;103(2):151423. doi: 10.1016/j.ejcb.2024.151423. PMID: 38659736 Free PMC article. Updated. Preprint.
-
Native cyclase-associated protein and actin from Xenopus laevis oocytes form a unique 4:4 complex with a tripartite structure.J Biol Chem. 2021 Jan-Jun;296:100649. doi: 10.1016/j.jbc.2021.100649. Epub 2021 Apr 9. J Biol Chem. 2021. PMID: 33839148 Free PMC article.
-
Mechanisms of actin disassembly and turnover.J Cell Biol. 2023 Dec 4;222(12):e202309021. doi: 10.1083/jcb.202309021. Epub 2023 Nov 10. J Cell Biol. 2023. PMID: 37948068 Free PMC article. Review.
-
Lessons from yeast for clathrin-mediated endocytosis.Nat Cell Biol. 2011 Dec 22;14(1):2-10. doi: 10.1038/ncb2403. Nat Cell Biol. 2011. PMID: 22193158 Free PMC article. Review.
References
-
- Balcer HI, Goodman AL, Rodal AA, Smith E, Kugler J, Heuser JE, Goode BL. Coordinated regulation of actin filament turnover by a high-molecular weight Srv2/CAP complex, cofilin, profilin, and Aip1. Current Biology. 2003;13:2159–2169. - PubMed
-
- Bertling E, Quintero-Monzon O, Mattila PK, Goode BL, Lappalainen P. Mechanism and biological role of profilin-Srv2/CAP interaction. Journal of Cell Science. 2007;120:1225–1234. - PubMed
-
- Blanchoin L, Pollard TD. Interaction of actin monomers with Acanthamoeba actophorin (ADF/cofilin) and profilin. J Biol Chem. 1998;273:25106–25111. - PubMed
-
- Bosch M, Le K, Bugyi B, Correia J, Renault L, Carlier MF. Analysis of the function of Spire in actin assembly and its synergy with formin and profilin. Molecular Cell. 2007;28:555–568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
