

BBA52 facilitates Borrelia burgdorferi transmission from feeding ticks to murine hosts
- PMID: 20170377
- PMCID: PMC2832101
- DOI: 10.1086/651172
BBA52 facilitates Borrelia burgdorferi transmission from feeding ticks to murine hosts
Abstract
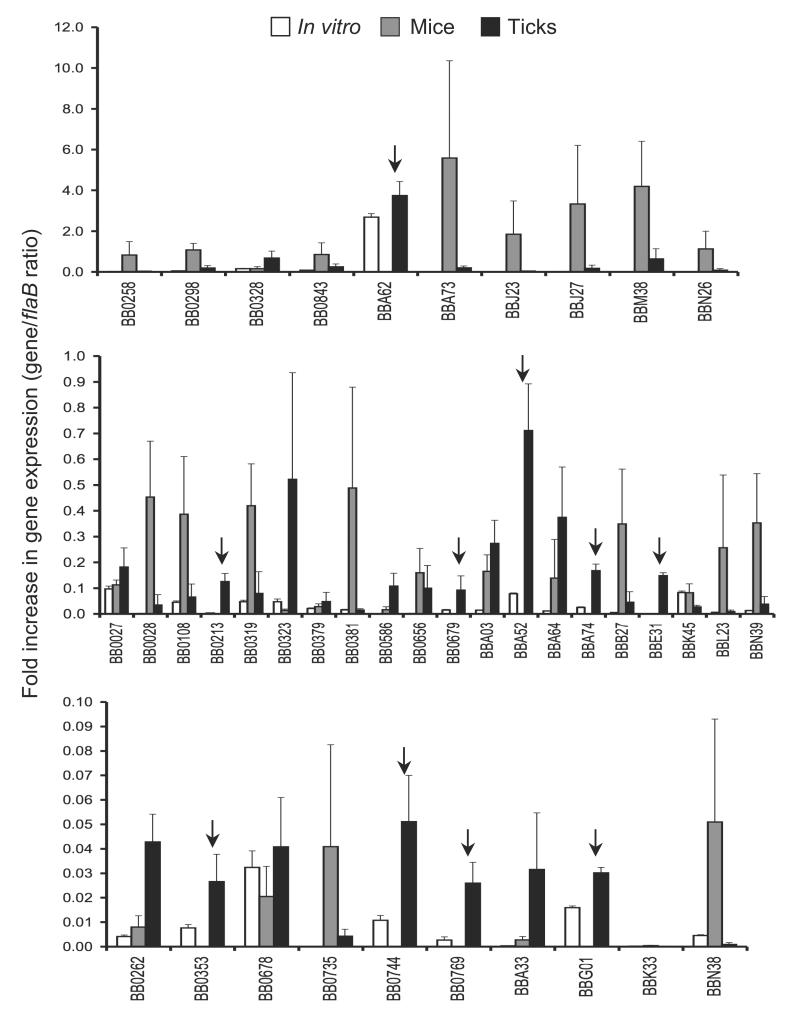
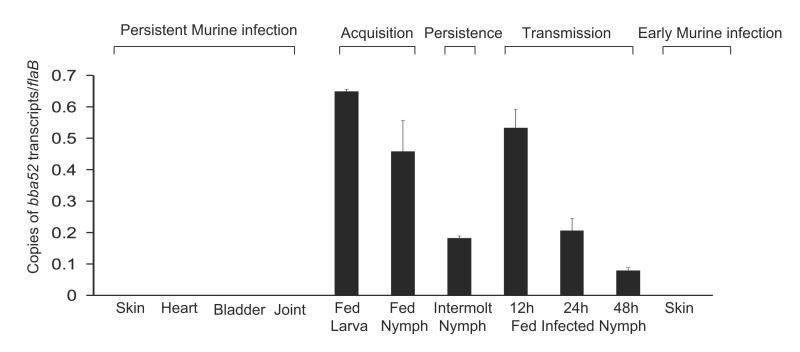
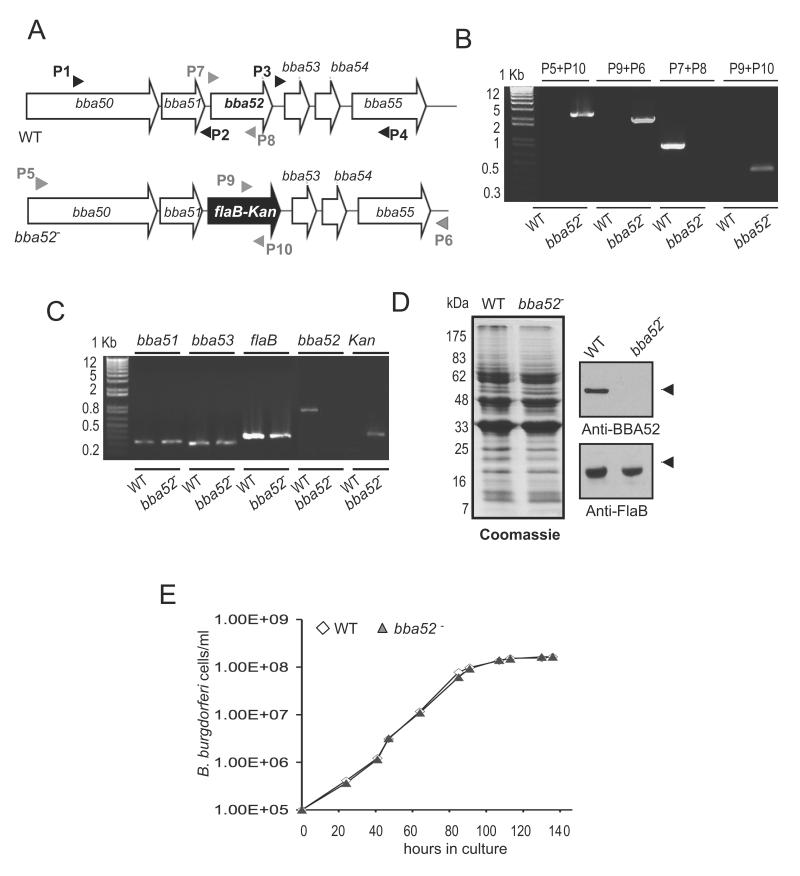
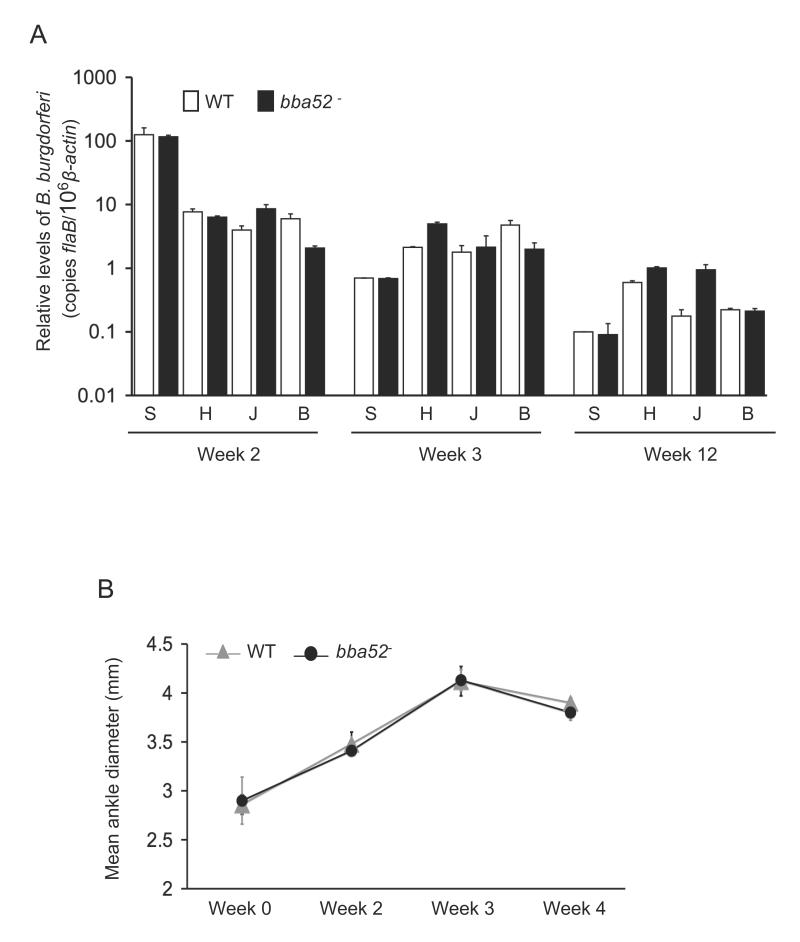
Borrelia burgdorferi, the pathogen of Lyme borreliosis, persists in nature through a tick-rodent transmission cycle. A selective assessment of the microbial transcriptome, limited to gene-encoding putative membrane proteins, reveals that bba52 transcription in vivo is strictly confined to the vector-specific portion of the microbial life cycle, with the highest levels of expression noted in feeding ticks and with swift down-regulation noted in mice. bba52 deletion did not affect murine disease as assessed by the genesis of arthritis and carditis or long-term persistence of pathogens in mice or ticks. However, bba52 deficiency did impair microbial transitions between hosts and vector, defects that could be fully rescued when bba52 expression was genetically restored to the original genomic locus. These studies establish that BBA52 facilitates vector-host transitions by the pathogen and therefore is a potential antigenic target for interference with transmission of B. burgdorferi from ticks to mammalian hosts.
Figures
Similar articles
-
A differential role for BB0365 in the persistence of Borrelia burgdorferi in mice and ticks.J Infect Dis. 2008 Jan 1;197(1):148-55. doi: 10.1086/523764. J Infect Dis. 2008. PMID: 18171298
-
Identification of Lyme borreliae proteins promoting vertebrate host blood-specific spirochete survival in Ixodes scapularis nymphs using artificial feeding chambers.Ticks Tick Borne Dis. 2018 Jul;9(5):1057-1063. doi: 10.1016/j.ttbdis.2018.03.033. Epub 2018 Apr 4. Ticks Tick Borne Dis. 2018. PMID: 29653905 Free PMC article.
-
Borrelia burgdorferi BBA52 is a potential target for transmission blocking Lyme disease vaccine.Vaccine. 2011 Nov 8;29(48):9012-9. doi: 10.1016/j.vaccine.2011.09.035. Epub 2011 Sep 21. Vaccine. 2011. PMID: 21945261 Free PMC article.
-
Borrelia burgdorferi protein interactions critical for microbial persistence in mammals.Cell Microbiol. 2019 Feb;21(2):e12885. doi: 10.1111/cmi.12885. Epub 2018 Jul 8. Cell Microbiol. 2019. PMID: 29934966 Free PMC article. Review.
-
New Insights Into CRASP-Mediated Complement Evasion in the Lyme Disease Enzootic Cycle.Front Cell Infect Microbiol. 2020 Jan 30;10:1. doi: 10.3389/fcimb.2020.00001. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32083019 Free PMC article. Review.
Cited by
-
Borrelia burgdorferi needs chemotaxis to establish infection in mammals and to accomplish its enzootic cycle.Infect Immun. 2012 Jul;80(7):2485-92. doi: 10.1128/IAI.00145-12. Epub 2012 Apr 16. Infect Immun. 2012. PMID: 22508862 Free PMC article.
-
Role of the surface lipoprotein BBA07 in the enzootic cycle of Borrelia burgdorferi.Infect Immun. 2010 Jul;78(7):2910-8. doi: 10.1128/IAI.00372-10. Epub 2010 Apr 26. Infect Immun. 2010. PMID: 20421380 Free PMC article.
-
A surface enolase participates in Borrelia burgdorferi-plasminogen interaction and contributes to pathogen survival within feeding ticks.Infect Immun. 2012 Jan;80(1):82-90. doi: 10.1128/IAI.05671-11. Epub 2011 Oct 24. Infect Immun. 2012. PMID: 22025510 Free PMC article.
-
Borrelia burgdorferi linear plasmid 38 is dispensable for completion of the mouse-tick infectious cycle.Infect Immun. 2011 Sep;79(9):3510-7. doi: 10.1128/IAI.05014-11. Epub 2011 Jun 27. Infect Immun. 2011. PMID: 21708994 Free PMC article.
-
The BBA33 lipoprotein binds collagen and impacts Borrelia burgdorferi pathogenesis.Mol Microbiol. 2015 Apr;96(1):68-83. doi: 10.1111/mmi.12921. Epub 2015 Jan 30. Mol Microbiol. 2015. PMID: 25560615 Free PMC article.
References
-
- Piesman J, Eisen L. Prevention of Tick-Borne Diseases. Annu Rev Entomol. 2008;53:323–343. - PubMed
-
- Barthold SW, DeSouza M, Fikrig E, Persing DH. Lyme borreliosis in the laboratory mouse. In: Schuster SE, editor. Lyme disease. Cold Spring Harbor Laboratory; Cold Spring Harbor, NY: 1992. pp. 223–242.
-
- Casjens S, Palmer N, van Vugt R, et al. A bacterial genome in flux: the twelve linear and nine circular extrachromosomal DNAs in an infectious isolate of the Lyme disease spirochete Borrelia burgdorferi. Mol Microbiol. 2000;35:490–516. - PubMed
-
- Fraser CM, Casjens S, Huang WM, et al. Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature. 1997;390:580–6. - PubMed
