

Identification and characterization of high molecular weight complexes formed by matrix AAA proteases and prohibitins in mitochondria of Arabidopsis thaliana
- PMID: 20172857
- PMCID: PMC2857092
- DOI: 10.1074/jbc.M109.063644
Identification and characterization of high molecular weight complexes formed by matrix AAA proteases and prohibitins in mitochondria of Arabidopsis thaliana
Abstract
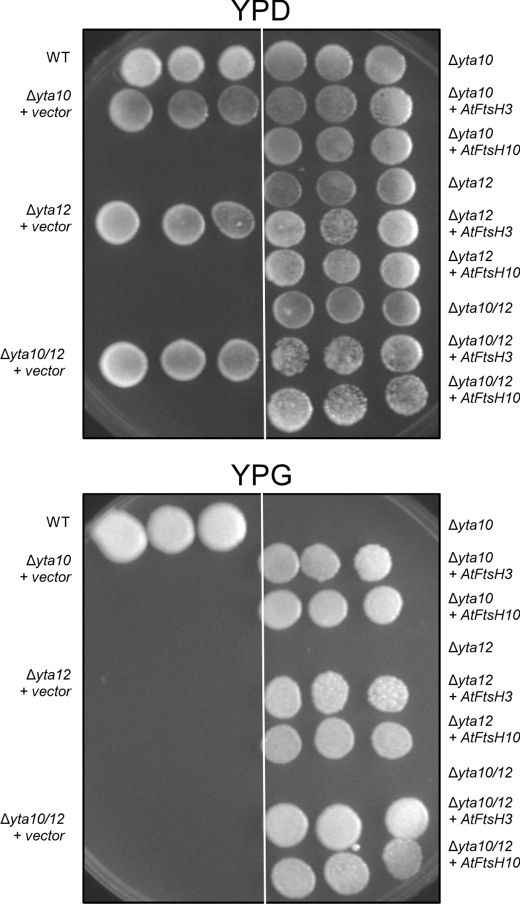
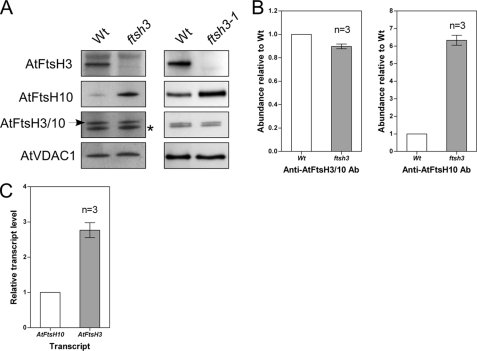
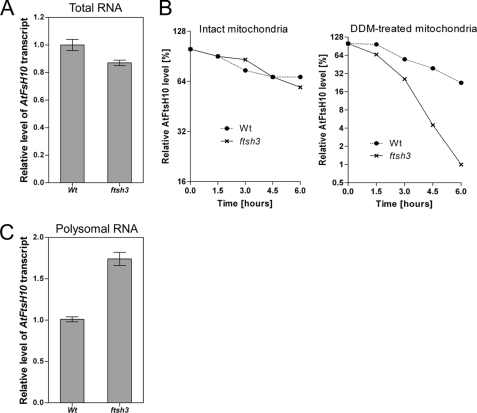
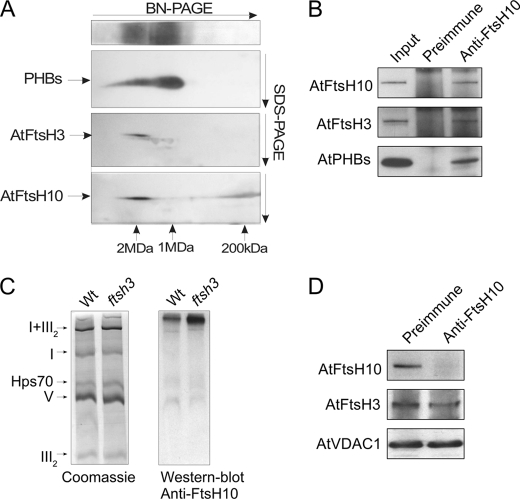
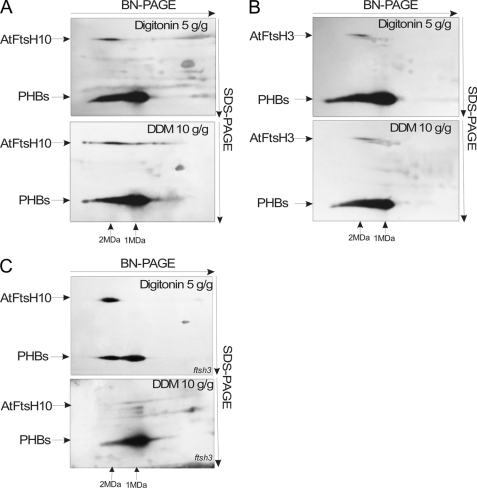
We identify and characterize two matrix (m)-AAA proteases (AtFtsH3 and AtFtsH10) present in the mitochondria of Arabidopsis thaliana. AtFtsH3 is the predominant protease in leaves of wild type plants. Both proteases assemble with prohibitins (PHBs) into high molecular weight complexes (approximately 2 MDa), similarly to their yeast counterparts. A smaller PHB complex (approximately 1 MDa), without the m-AAA proteases, was also detected. Unlike in yeast, stable prohibitin-independent high molecular weight assemblies of m-AAA proteases could not be identified in A. thaliana. AtFtsH3 and AtFtsH10 form at least two types of m-AAA-PHB complexes in wild type plants. The one type contains PHBs and AtFtsH3, and the second one is composed of PHBs and both AtFtsH3 and AtFtsH10. Complexes composed of PHBs and AtFtsH10 were found in an Arabidopsis mutant lacking AtFtsH3 (ftsh3). Thus, both AtFtsH3 and AtFtsH10 may form hetero- and homo-oligomeric complexes with prohibitins. The increased level of AtFtsH10 observed in ftsh3 suggests that functions of the homo- and hetero-oligomeric complexes containing AtFtsH3 can be at least partially substituted by AtFtsH10 homo-oligomers. The steady-state level of the AtFtsH10 transcripts did not change in ftsh3 compared with wild type plants, but we found that almost twice more of the AtFtsH10 transcripts were associated with polysomes in ftsh3. Based on this result, we assume that the AtFtsH10 protein is synthesized at a higher rate in the ftsh3 mutant. Our results provide the first data on the composition of m-AAA and PHB complexes in plant mitochondria and suggest that the abundance of m-AAA proteases is regulated not only at the transcriptional but also at the translational level.
Figures
Similar articles
-
m-AAA Complexes Are Not Crucial for the Survival of Arabidopsis Under Optimal Growth Conditions Despite Their Importance for Mitochondrial Translation.Plant Cell Physiol. 2018 May 1;59(5):1006-1016. doi: 10.1093/pcp/pcy041. Plant Cell Physiol. 2018. PMID: 29462458
-
Plant mitochondria contain at least two i-AAA-like complexes.Plant Mol Biol. 2005 Sep;59(2):239-52. doi: 10.1007/s11103-005-8766-3. Plant Mol Biol. 2005. PMID: 16247555
-
ATP-dependent proteases in biogenesis and maintenance of plant mitochondria.Biochim Biophys Acta. 2010 Jun-Jul;1797(6-7):1071-5. doi: 10.1016/j.bbabio.2010.02.027. Epub 2010 Mar 1. Biochim Biophys Acta. 2010. PMID: 20193658 Review.
-
Unraveling the functions of type II-prohibitins in Arabidopsis mitochondria.Plant Mol Biol. 2015 Jun;88(3):249-67. doi: 10.1007/s11103-015-0320-3. Epub 2015 Apr 21. Plant Mol Biol. 2015. PMID: 25896400
-
m-AAA proteases, mitochondrial calcium homeostasis and neurodegeneration.Cell Res. 2018 Mar;28(3):296-306. doi: 10.1038/cr.2018.17. Epub 2018 Feb 16. Cell Res. 2018. PMID: 29451229 Free PMC article. Review.
Cited by
-
Lack of FTSH4 Protease Affects Protein Carbonylation, Mitochondrial Morphology, and Phospholipid Content in Mitochondria of Arabidopsis: New Insights into a Complex Interplay.Plant Physiol. 2016 Aug;171(4):2516-35. doi: 10.1104/pp.16.00370. Epub 2016 Jun 13. Plant Physiol. 2016. PMID: 27297677 Free PMC article.
-
The mitochondrial AAA protease FTSH3 regulates Complex I abundance by promoting its disassembly.Plant Physiol. 2021 May 27;186(1):599-610. doi: 10.1093/plphys/kiab074. Plant Physiol. 2021. PMID: 33616659 Free PMC article.
-
AAA Proteases: Guardians of Mitochondrial Function and Homeostasis.Cells. 2018 Oct 11;7(10):163. doi: 10.3390/cells7100163. Cells. 2018. PMID: 30314276 Free PMC article. Review.
-
The interplay of light and oxygen in the reactive oxygen stress response of Chlamydomonas reinhardtii dissected by quantitative mass spectrometry.Mol Cell Proteomics. 2014 Apr;13(4):969-89. doi: 10.1074/mcp.M113.032771. Epub 2014 Jan 29. Mol Cell Proteomics. 2014. PMID: 24482124 Free PMC article.
-
Protein Processing in Plant Mitochondria Compared to Yeast and Mammals.Front Plant Sci. 2022 Feb 2;13:824080. doi: 10.3389/fpls.2022.824080. eCollection 2022. Front Plant Sci. 2022. PMID: 35185991 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
