

Highly differentiated, resting gn-specific memory CD8+ T cells persist years after infection by andes hantavirus
- PMID: 20174562
- PMCID: PMC2824805
- DOI: 10.1371/journal.ppat.1000779
Highly differentiated, resting gn-specific memory CD8+ T cells persist years after infection by andes hantavirus
Abstract
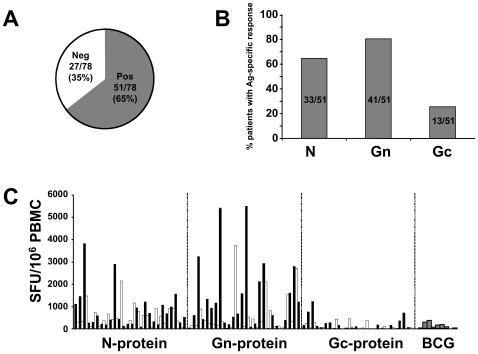
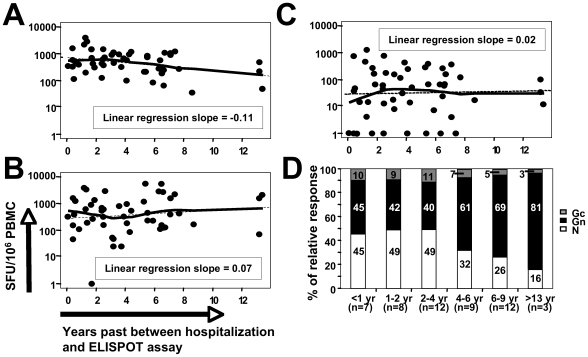
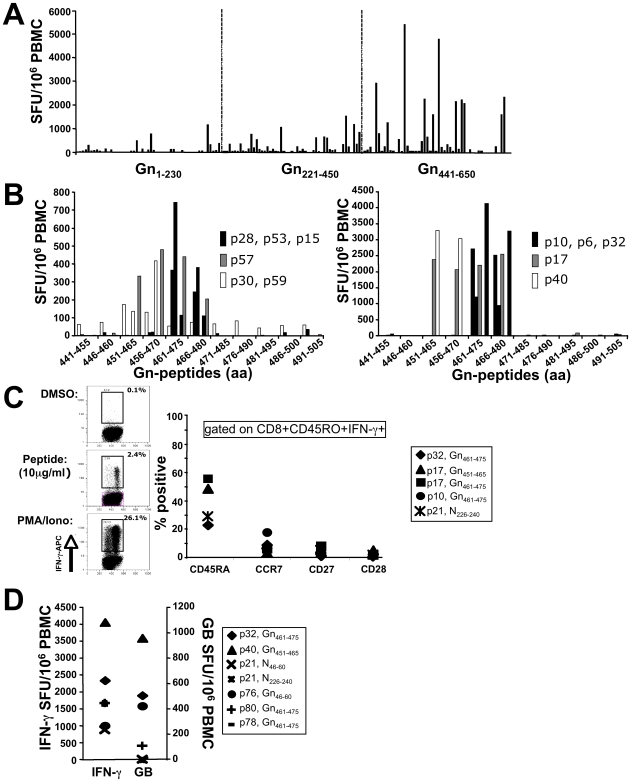
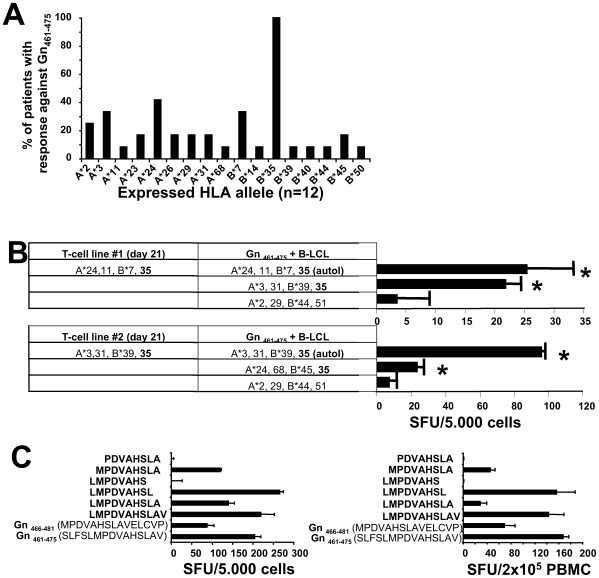
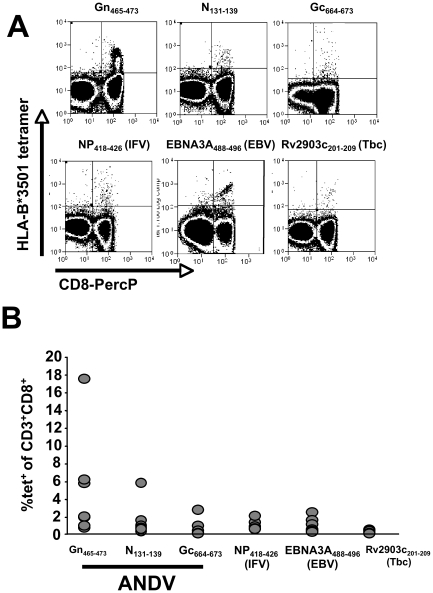
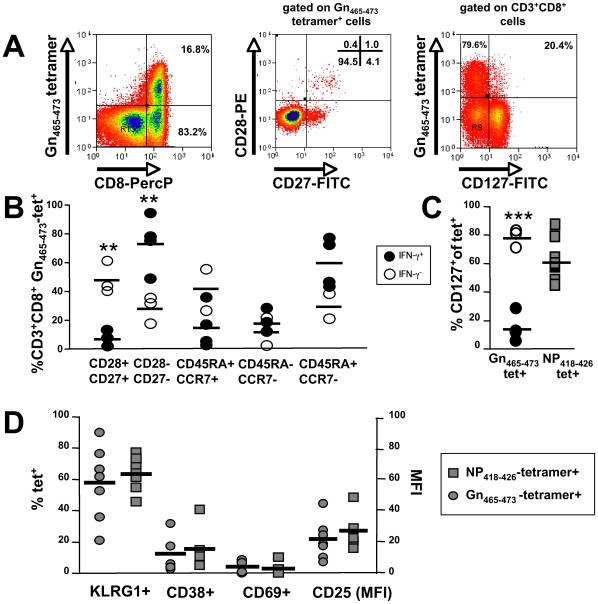
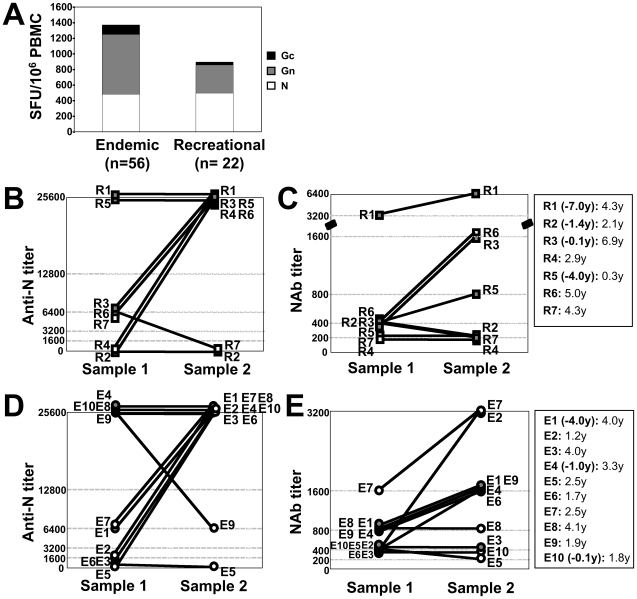
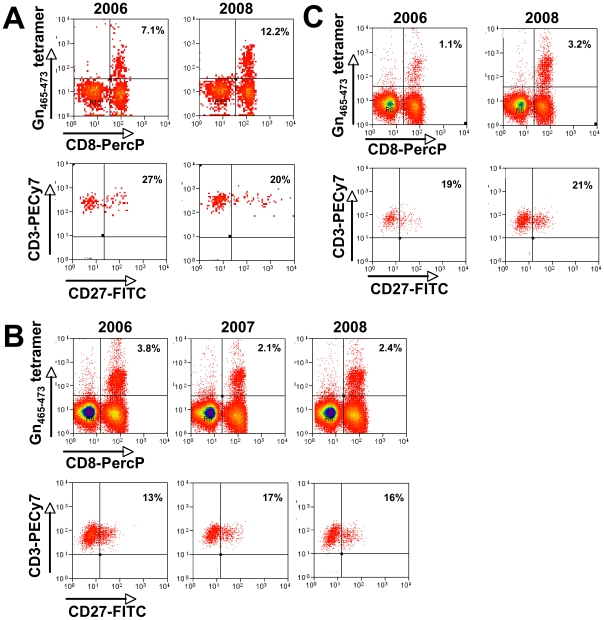
In man, infection with South American Andes virus (ANDV) causes hantavirus cardiopulmonary syndrome (HCPS). HCPS due to ANDV is endemic in Southern Chile and much of Argentina and increasing numbers of cases are reported all over South America. A case-fatality rate of about 36% together with the absence of successful antiviral therapies urge the development of a vaccine. Although T-cell responses were shown to be critically involved in immunity to hantaviruses in mouse models, no data are available on the magnitude, specificity and longevity of ANDV-specific memory T-cell responses in patients. Using sets of overlapping peptides in IFN-gamma ELISPOT assays, we herein show in 78 Chilean convalescent patients that Gn-derived epitopes were immunodominant as compared to those from the N- and Gc-proteins. Furthermore, while the relative contribution of the N-specific response significantly declined over time, Gn-specific responses remained readily detectable ex vivo up to 13 years after the acute infection. Tetramer analysis further showed that up to 16.8% of all circulating CD3(+)CD8(+) T cells were specific for the single HLA-B*3501-restricted epitope Gn(465-473) years after the acute infection. Remarkably, Gn(465-473)-specific cells readily secreted IFN-gamma, granzyme B and TNF-alpha but not IL-2 upon stimulation and showed a 'revertant' CD45RA(+)CD27(-)CD28(-)CCR7(-)CD127(-) effector memory phenotype, thereby resembling a phenotype seen in other latent virus infections. Most intriguingly, titers of neutralizing antibodies increased over time in 10/17 individuals months to years after the acute infection and independently of whether they were residents of endemic areas or not. Thus, our data suggest intrinsic, latent antigenic stimulation of Gn-specific T-cells. However, it remains a major task for future studies to proof this hypothesis by determination of viral antigen in convalescent patients. Furthermore, it remains to be seen whether Gn-specific T cells are critical for viral control and protective immunity. If so, Gn-derived immunodominant epitopes could be of high value for future ANDV vaccines.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Weissenbacher MC, Cura E, Segura EL, Hortal M, Baek LJ, et al. Serological evidence of human Hantavirus infection in Argentina, Bolivia and Uruguay. Medicina (B Aires) 1996;56:17–22. - PubMed
-
- da Silva MV, Vasconcelos MJ, Hidalgo NT, Veiga AP, Canzian M, et al. Hantavirus pulmonary syndrome. Report of the first three cases in Sao Paulo, Brazil. Rev Inst Med Trop Sao Paulo. 1997;39:231–234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
