

Arterivirus Nsp1 modulates the accumulation of minus-strand templates to control the relative abundance of viral mRNAs
- PMID: 20174607
- PMCID: PMC2824749
- DOI: 10.1371/journal.ppat.1000772
Arterivirus Nsp1 modulates the accumulation of minus-strand templates to control the relative abundance of viral mRNAs
Abstract
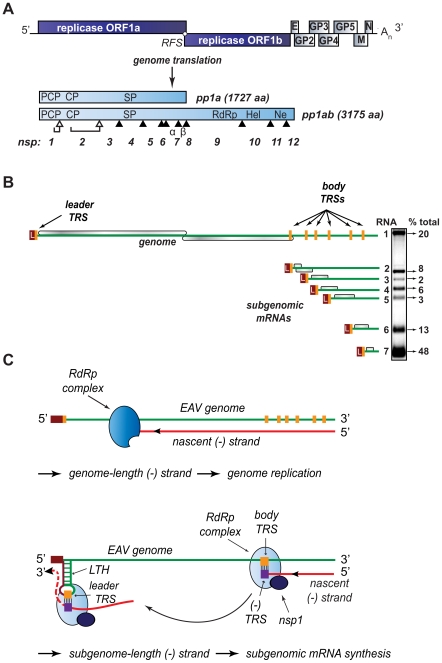
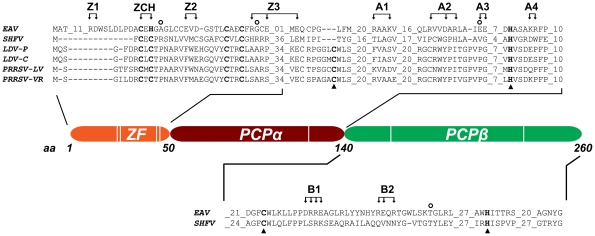
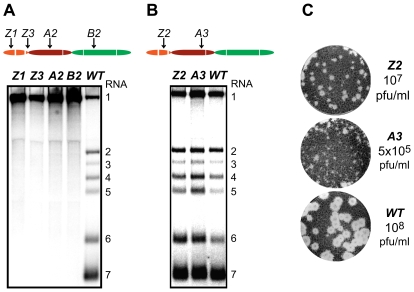
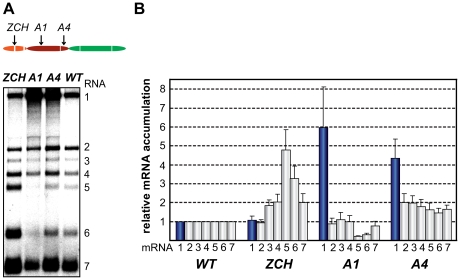
The gene expression of plus-strand RNA viruses with a polycistronic genome depends on translation and replication of the genomic mRNA, as well as synthesis of subgenomic (sg) mRNAs. Arteriviruses and coronaviruses, distantly related members of the nidovirus order, employ a unique mechanism of discontinuous minus-strand RNA synthesis to generate subgenome-length templates for the synthesis of a nested set of sg mRNAs. Non-structural protein 1 (nsp1) of the arterivirus equine arteritis virus (EAV), a multifunctional regulator of viral RNA synthesis and virion biogenesis, was previously implicated in controlling the balance between genome replication and sg mRNA synthesis. Here, we employed reverse and forward genetics to gain insight into the multiple regulatory roles of nsp1. Our analysis revealed that the relative abundance of viral mRNAs is tightly controlled by an intricate network of interactions involving all nsp1 subdomains. Distinct nsp1 mutations affected the quantitative balance among viral mRNA species, and our data implicate nsp1 in controlling the accumulation of full-length and subgenome-length minus-strand templates for viral mRNA synthesis. The moderate differential changes in viral mRNA abundance of nsp1 mutants resulted in similarly altered viral protein levels, but progeny virus yields were greatly reduced. Pseudorevertant analysis provided compelling genetic evidence that balanced EAV mRNA accumulation is critical for efficient virus production. This first report on protein-mediated, mRNA-specific control of nidovirus RNA synthesis reveals the existence of an integral control mechanism to fine-tune replication, sg mRNA synthesis, and virus production, and establishes a major role for nsp1 in coordinating the arterivirus replicative cycle.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Arterivirus subgenomic mRNA synthesis and virion biogenesis depend on the multifunctional nsp1 autoprotease.J Virol. 2007 Oct;81(19):10496-505. doi: 10.1128/JVI.00683-07. Epub 2007 Jul 11. J Virol. 2007. PMID: 17626105 Free PMC article.
-
Genetic manipulation of arterivirus alternative mRNA leader-body junction sites reveals tight regulation of structural protein expression.J Virol. 2000 Dec;74(24):11642-53. doi: 10.1128/jvi.74.24.11642-11653.2000. J Virol. 2000. PMID: 11090163 Free PMC article.
-
Adaptive Mutations in Replicase Transmembrane Subunits Can Counteract Inhibition of Equine Arteritis Virus RNA Synthesis by Cyclophilin Inhibitors.J Virol. 2019 Aug 28;93(18):e00490-19. doi: 10.1128/JVI.00490-19. Print 2019 Sep 15. J Virol. 2019. PMID: 31243130 Free PMC article.
-
Nidovirus RNA polymerases: Complex enzymes handling exceptional RNA genomes.Virus Res. 2017 Apr 15;234:58-73. doi: 10.1016/j.virusres.2017.01.023. Epub 2017 Feb 6. Virus Res. 2017. PMID: 28174054 Free PMC article. Review.
-
Nidovirus transcription: how to make sense...?J Gen Virol. 2006 Jun;87(Pt 6):1403-1421. doi: 10.1099/vir.0.81611-0. J Gen Virol. 2006. PMID: 16690906 Review.
Cited by
-
The footprint of genome architecture in the largest genome expansion in RNA viruses.PLoS Pathog. 2013;9(7):e1003500. doi: 10.1371/journal.ppat.1003500. Epub 2013 Jul 18. PLoS Pathog. 2013. PMID: 23874204 Free PMC article.
-
The dimeric conformation of PRRSV nsp1α is important for its ability to regulate viral RNA synthesis.Vet Res. 2025 May 21;56(1):105. doi: 10.1186/s13567-025-01537-5. Vet Res. 2025. PMID: 40399953 Free PMC article.
-
Common and unique features of viral RNA-dependent polymerases.Cell Mol Life Sci. 2014 Nov;71(22):4403-20. doi: 10.1007/s00018-014-1695-z. Epub 2014 Aug 1. Cell Mol Life Sci. 2014. PMID: 25080879 Free PMC article. Review.
-
An intra-family conserved high-order RNA structure within the M ORF is important for arterivirus subgenomic RNA accumulation and infectious virus production.J Virol. 2025 May 20;99(5):e0216724. doi: 10.1128/jvi.02167-24. Epub 2025 Apr 7. J Virol. 2025. PMID: 40192289 Free PMC article.
-
Arterivirus nsp12 versus the coronavirus nsp16 2'-O-methyltransferase: comparison of the C-terminal cleavage products of two nidovirus pp1ab polyproteins.J Gen Virol. 2015 Sep;96(9):2643-2655. doi: 10.1099/vir.0.000209. Epub 2015 Jun 3. J Gen Virol. 2015. PMID: 26041874 Free PMC article.
References
-
- Miller WA, Koev G. Synthesis of subgenomic RNAs by positive-strand RNA viruses. Virology. 2000;273:1–8. - PubMed
-
- Molenkamp R, van Tol H, Rozier BCD, van der Meer Y, Spaan WJM, Snijder EJ. The arterivirus replicase is the only viral protein required for genome replication and subgenomic mRNA transcription. J Gen Virol. 2000;81:2491–2496. - PubMed
-
- Sawicki SG, Sawicki DL. Coronaviruses use discontinuous extension for synthesis of subgenome-length negative strands. Adv Exp Med Biol. 1995;380:499–506. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
