

A rearrangement of the guanosine-binding site establishes an extended network of functional interactions in the Tetrahymena group I ribozyme active site
- PMID: 20175542
- PMCID: PMC2860537
- DOI: 10.1021/bi902200n
A rearrangement of the guanosine-binding site establishes an extended network of functional interactions in the Tetrahymena group I ribozyme active site
Abstract
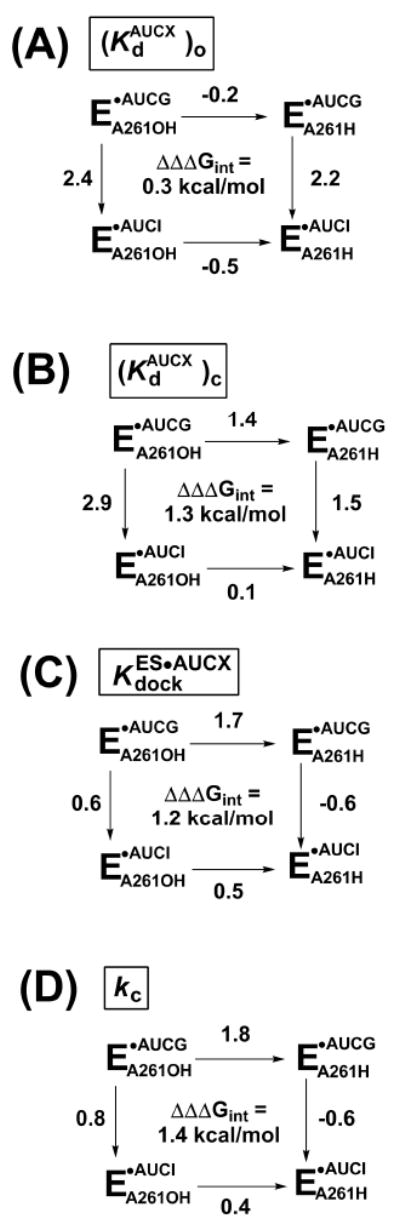
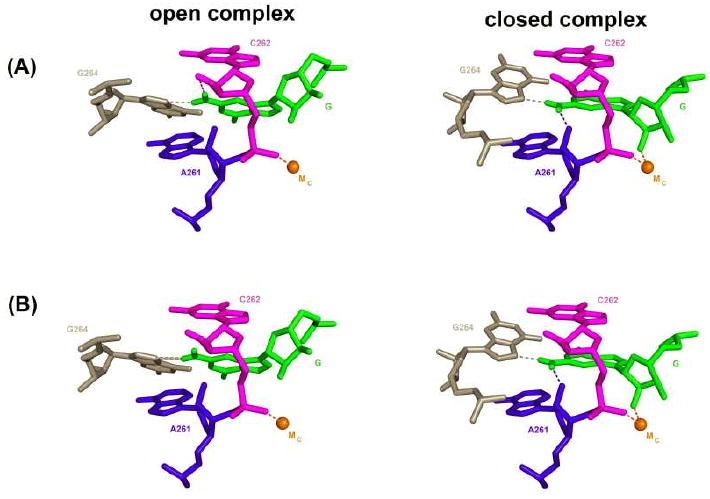
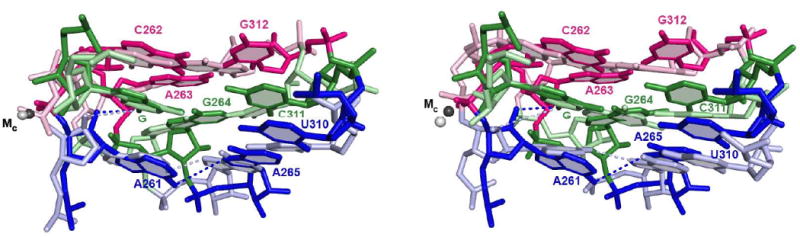
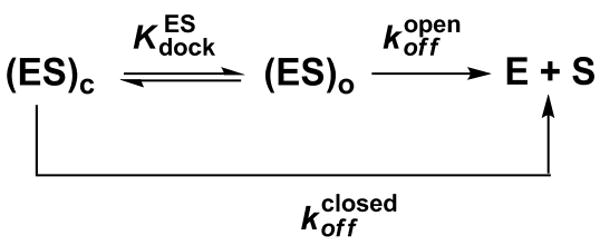
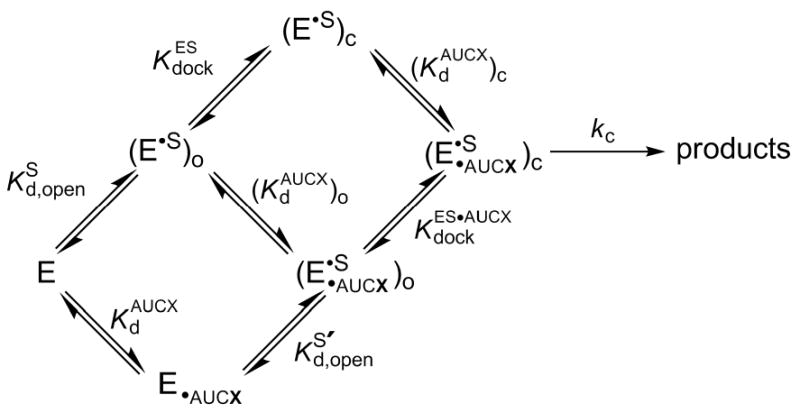
Protein enzymes appear to use extensive packing and hydrogen bonding interactions to precisely position catalytic groups within active sites. Because of their inherent backbone flexibility and limited side chain repertoire, RNA enzymes face additional challenges relative to proteins in precisely positioning substrates and catalytic groups. Here, we use the group I ribozyme to probe the existence, establishment, and functional consequences of an extended network of interactions in an RNA active site. The group I ribozyme catalyzes a site-specific attack of guanosine on an oligonucleotide substrate. We previously determined that the hydrogen bond between the exocyclic amino group of guanosine and the 2'-hydroxyl group at position A261 of the Tetrahymena group I ribozyme contributes to overall catalysis. We now use functional data, aided by double mutant cycles, to probe this hydrogen bond in the individual reaction steps of the catalytic cycle. Our results indicate that this hydrogen bond is not formed upon guanosine binding to the ribozyme but instead forms at a later stage of the catalytic cycle. Formation of this hydrogen bond is correlated with other structural rearrangements in the ribozyme's active site that are promoted by docking of the oligonucleotide substrate into the ribozyme's active site, and disruption of this interaction has deleterious consequences for the chemical transformation within the ternary complex. These results, combined with earlier results, provide insight into the nature of the multiple conformational steps used by the Tetrahymena group I ribozyme to achieve its active structure and reveal an intricate, extended network of interactions that is used to establish catalytic interactions within this RNA's active site.
Figures


References
-
- Jencks WP. Economics of enzyme catalysis. Cold Spring Harb Symp Quant Biol. 1987;52:65–73. - PubMed
-
- Ditzler MA, Aleman EA, Rueda D, Walter NG. Focus on function: single molecule RNA enzymology. Biopolymers. 2007;87:302–316. - PubMed
-
- Blount KF, Uhlenbeck OC. The structure-function dilemma of the hammerhead ribozyme. Annu Rev Biophys Biomol Struct. 2005;34:415–440. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
