

Animal HECT ubiquitin ligases: evolution and functional implications
- PMID: 20175895
- PMCID: PMC2837046
- DOI: 10.1186/1471-2148-10-56
Animal HECT ubiquitin ligases: evolution and functional implications
Abstract
Background: HECT ubiquitin ligases (HECT E3s) are key components of the eukaryotic ubiquitin-proteasome system and are involved in the genesis of several human diseases. In this study, I analyze the patterns of diversification of HECT E3s since animals emerged in order to provide the right framework to understand the functional data available for proteins of this family.
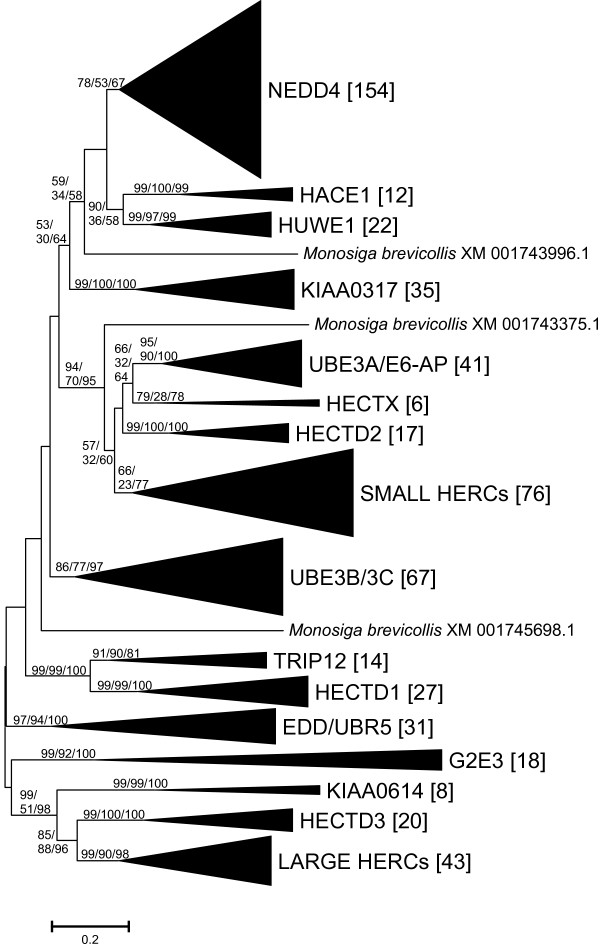
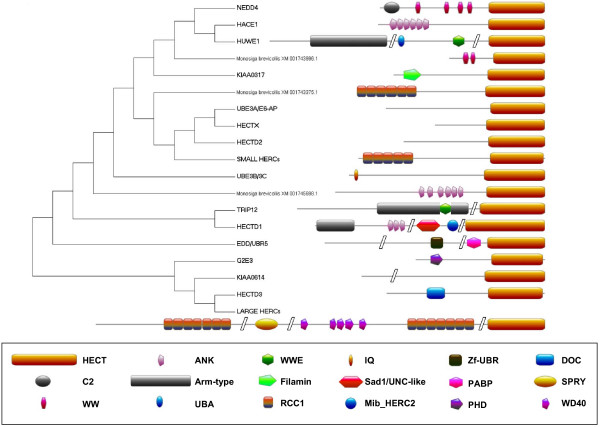
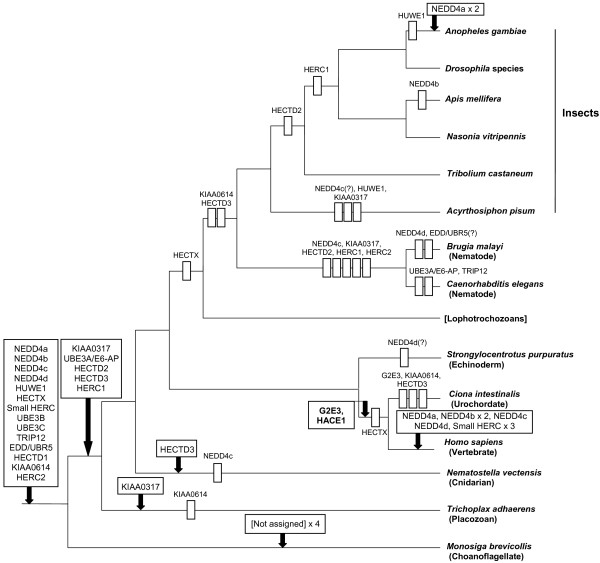
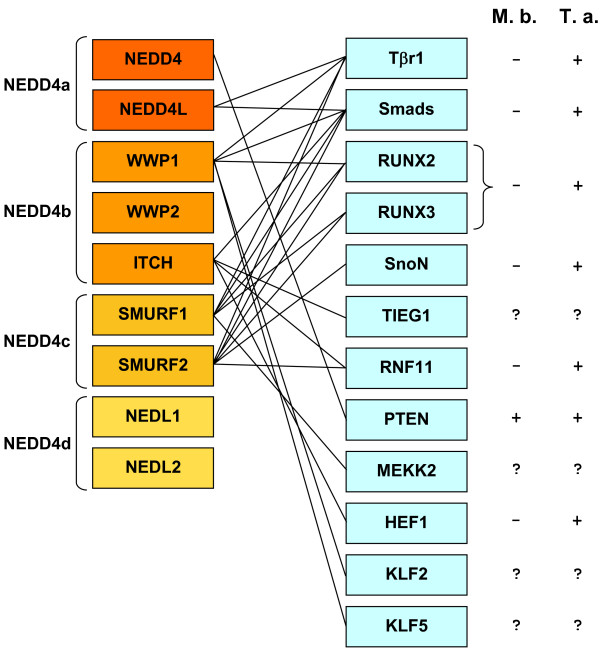
Results: I show that the current classification of HECT E3s into three groups (NEDD4-like E3s, HERCs and single-HECT E3s) is fundamentally incorrect. First, the existence of a "Single-HECT E3s" group is not supported by phylogenetic analyses. Second, the HERC proteins must be divided into two subfamilies (Large HERCs, Small HERCs) that are evolutionarily very distant, their structural similarity being due to convergence and not to a common origin. Sequence and structural analyses show that animal HECT E3s can be naturally classified into 16 subfamilies. Almost all of them appeared either before animals originated or in early animal evolution. More recently, multiple gene losses have occurred independently in some lineages (nematodes, insects, urochordates), the same groups that have also lost genes of another type of E3s (RBR family). Interestingly, the emergence of some animal HECT E3s precedes the origin of key cellular systems that they regulate (TGF-beta and EGF signal transduction pathways; p53 family of transcription factors) and it can be deduced that distantly related HECT proteins have been independently co-opted to perform similar roles. This may contribute to explain why distantly related HECT E3s are involved in the genesis of multiple types of cancer.
Conclusions: The complex evolutionary history of HECT ubiquitin ligases in animals has been deciphered. The most appropriate model animals to study them and new theoretical and experimental lines of research are suggested by these results.
Figures
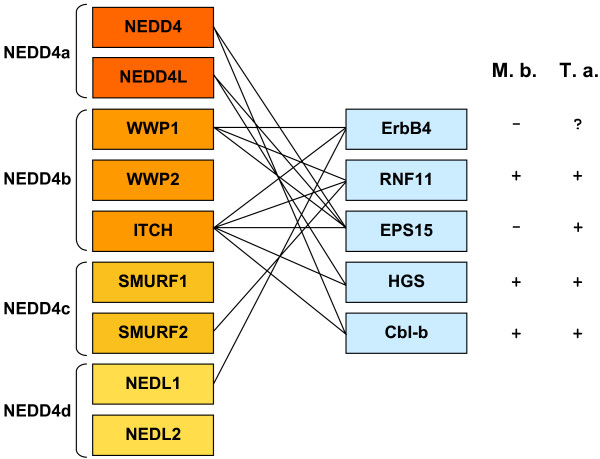
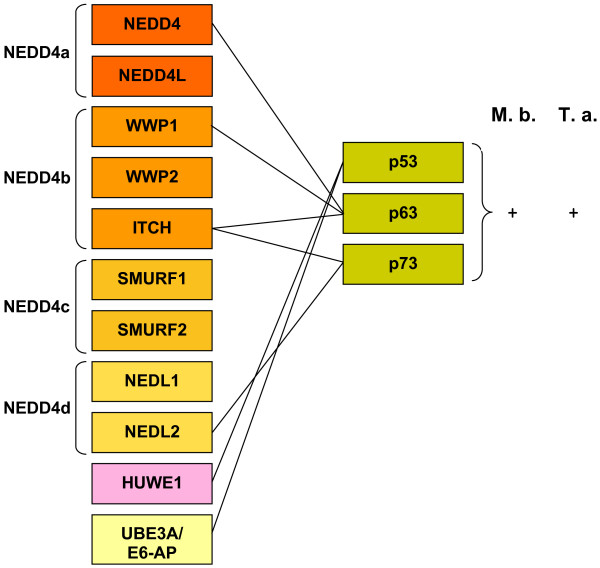
Similar articles
-
Origin and evolution of fungal HECT ubiquitin ligases.Sci Rep. 2018 Apr 23;8(1):6419. doi: 10.1038/s41598-018-24914-x. Sci Rep. 2018. PMID: 29686411 Free PMC article.
-
Evolution of plant HECT ubiquitin ligases.PLoS One. 2013 Jul 15;8(7):e68536. doi: 10.1371/journal.pone.0068536. Print 2013. PLoS One. 2013. PMID: 23869223 Free PMC article.
-
Adaptors as the regulators of HECT ubiquitin ligases.Cell Death Differ. 2021 Feb;28(2):455-472. doi: 10.1038/s41418-020-00707-6. Epub 2021 Jan 5. Cell Death Differ. 2021. PMID: 33402750 Free PMC article. Review.
-
β-Sheet Augmentation Is a Conserved Mechanism of Priming HECT E3 Ligases for Ubiquitin Ligation.J Mol Biol. 2018 Sep 14;430(18 Pt B):3218-3233. doi: 10.1016/j.jmb.2018.06.044. Epub 2018 Jun 28. J Mol Biol. 2018. PMID: 29964046
-
The HECT family of E3 ubiquitin ligases: multiple players in cancer development.Cancer Cell. 2008 Jul 8;14(1):10-21. doi: 10.1016/j.ccr.2008.06.001. Cancer Cell. 2008. PMID: 18598940 Review.
Cited by
-
A Smurf1 tale: function and regulation of an ubiquitin ligase in multiple cellular networks.Cell Mol Life Sci. 2013 Jul;70(13):2305-17. doi: 10.1007/s00018-012-1170-7. Epub 2012 Sep 25. Cell Mol Life Sci. 2013. PMID: 23007848 Free PMC article. Review.
-
Structural Basis for the Enzymatic Activity of the HACE1 HECT-Type E3 Ligase Through N-Terminal Helix Dimerization.Adv Sci (Weinh). 2023 Sep;10(27):e2207672. doi: 10.1002/advs.202207672. Epub 2023 Aug 3. Adv Sci (Weinh). 2023. PMID: 37537642 Free PMC article.
-
Early Origin and Evolution of the Angelman Syndrome Ubiquitin Ligase Gene Ube3a.Front Cell Neurosci. 2017 Mar 7;11:62. doi: 10.3389/fncel.2017.00062. eCollection 2017. Front Cell Neurosci. 2017. PMID: 28326016 Free PMC article. Review.
-
Comparative interactome analysis of α-arrestin families in human and Drosophila.Elife. 2024 Jan 25;12:RP88328. doi: 10.7554/eLife.88328. Elife. 2024. PMID: 38270169 Free PMC article.
-
Genome-wide identification, phylogenetic and expression analysis of the maize HECT E3 ubiquitin ligase genes.Genetica. 2019 Dec;147(5-6):391-400. doi: 10.1007/s10709-019-00080-4. Epub 2019 Nov 18. Genetica. 2019. PMID: 31741104
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
