

Structure and function of immunoglobulins
- PMID: 20176268
- PMCID: PMC3670108
- DOI: 10.1016/j.jaci.2009.09.046
Structure and function of immunoglobulins
Abstract
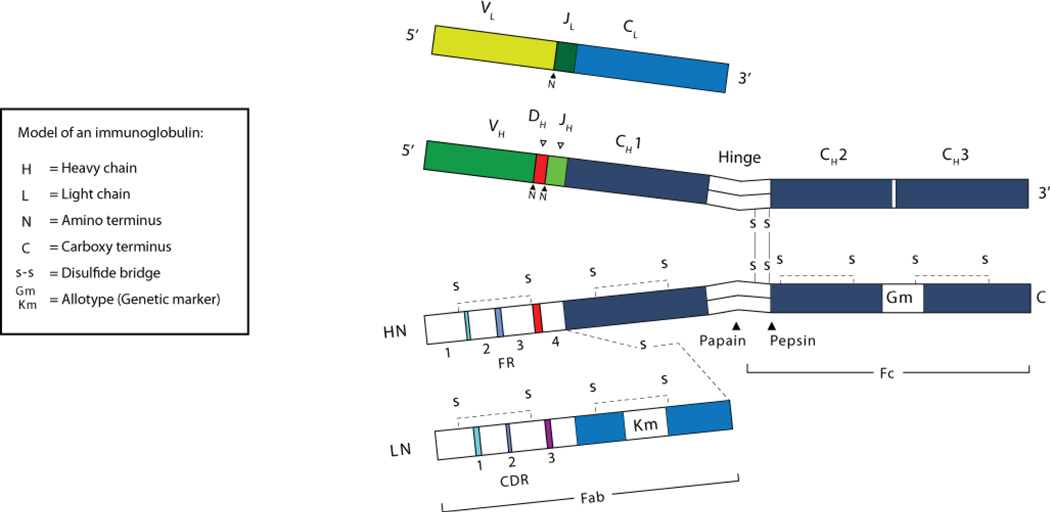
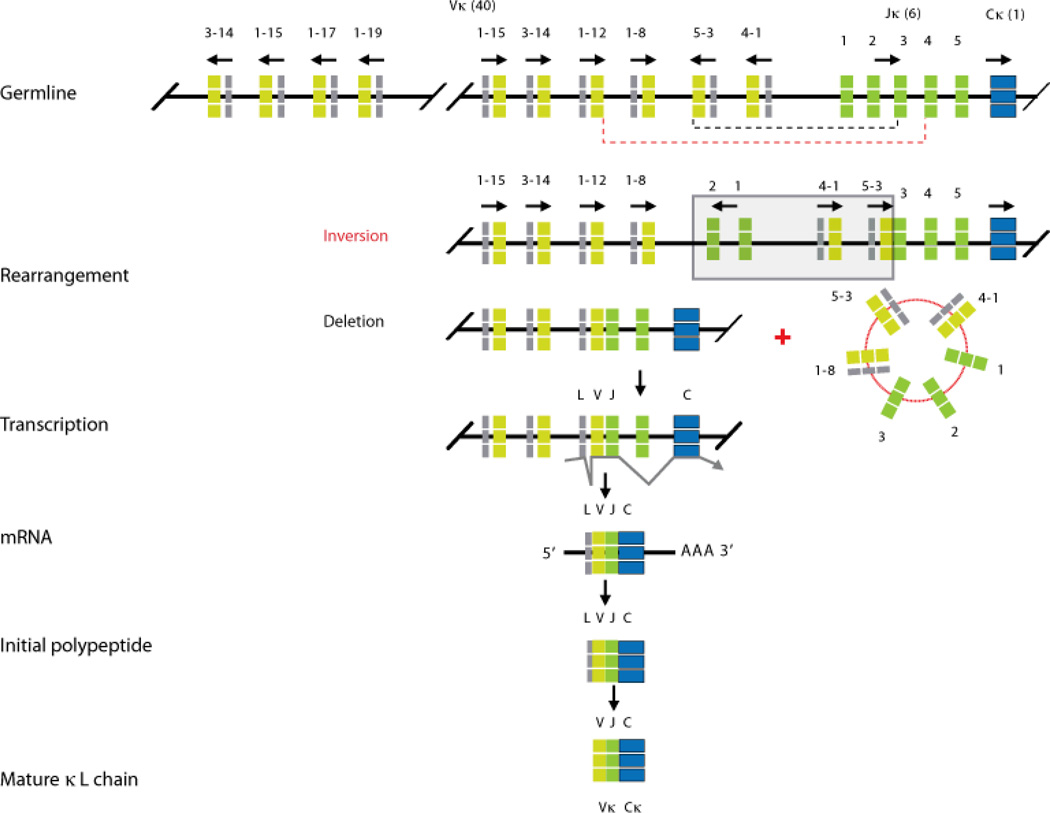
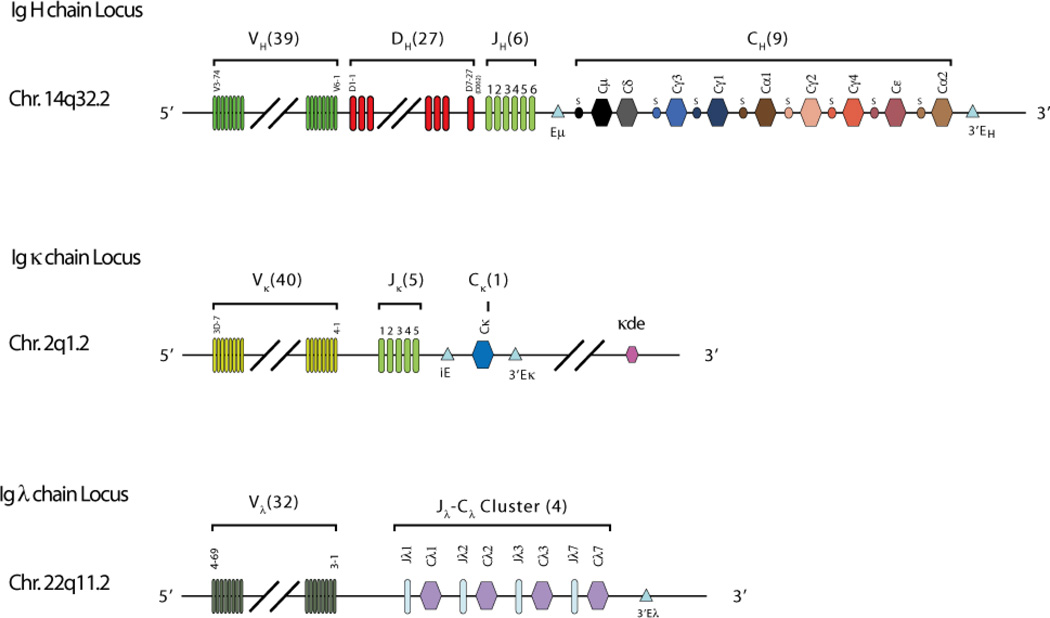
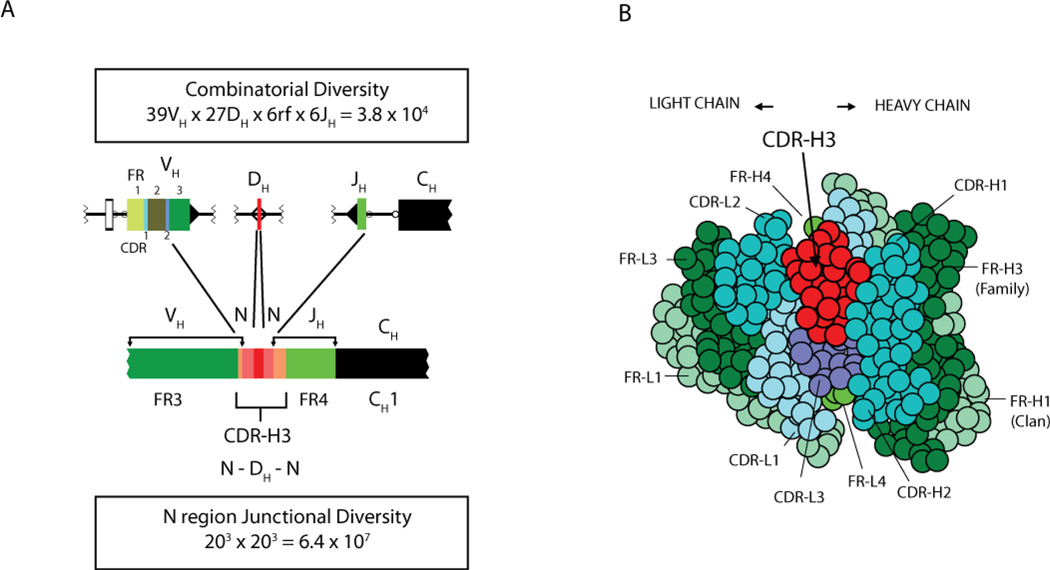
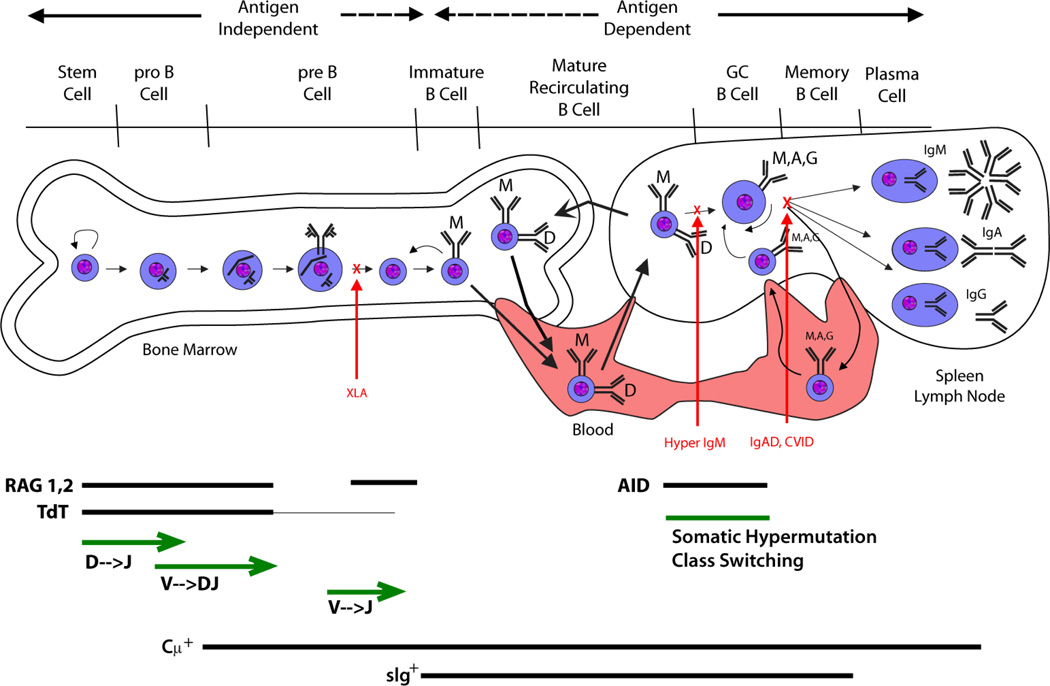
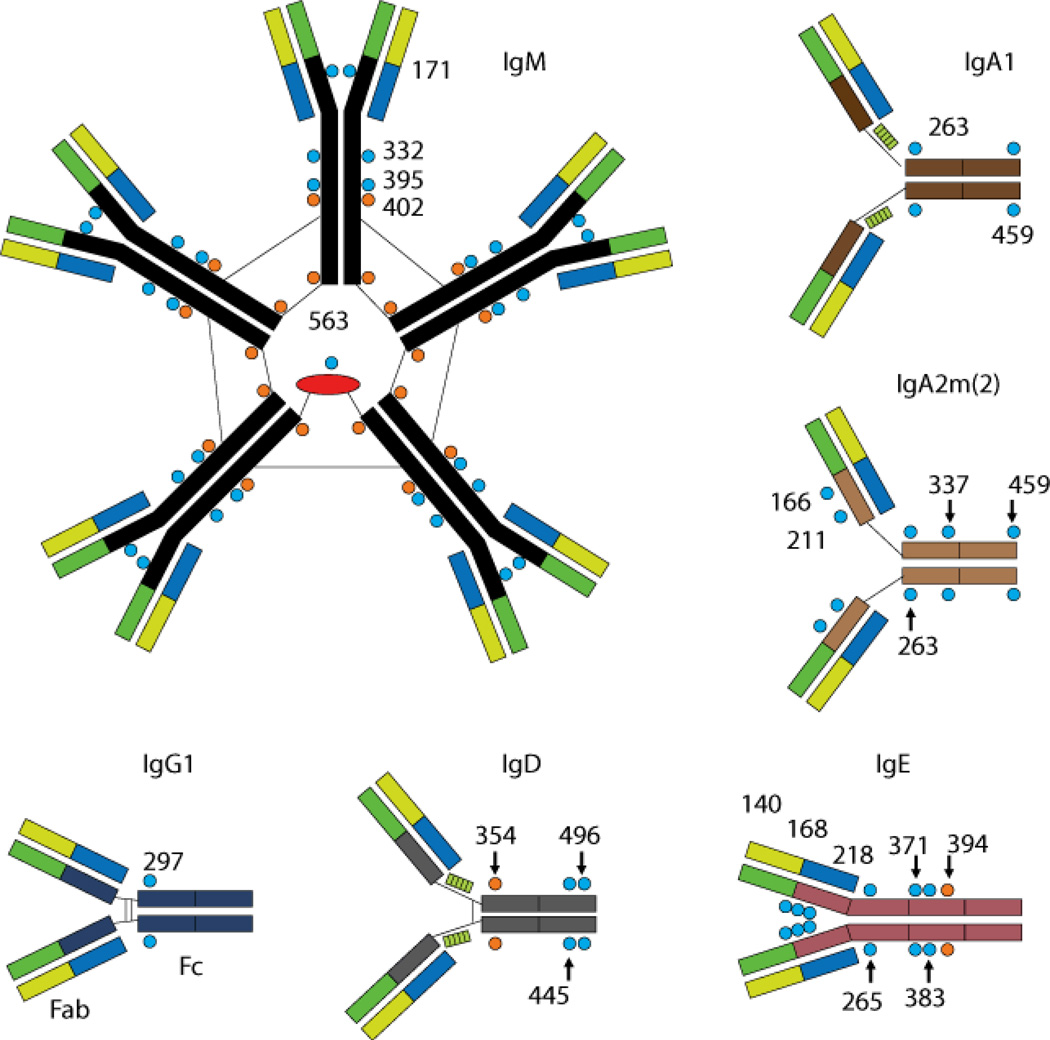
Immunoglobulins are heterodimeric proteins composed of 2 heavy and 2 light chains. They can be separated functionally into variable domains that bind antigens and constant domains that specify effector functions, such as activation of complement or binding to Fc receptors. The variable domains are created by means of a complex series of gene rearrangement events and can then be subjected to somatic hypermutation after exposure to antigen to allow affinity maturation. Each variable domain can be split into 3 regions of sequence variability termed the complementarity-determining regions (CDRs) and 4 regions of relatively constant sequence termed the framework regions. The 3 CDRs of the heavy chain are paired with the 3 CDRs of the light chain to form the antigen-binding site, as classically defined. The constant domains of the heavy chain can be switched to allow altered effector function while maintaining antigen specificity. There are 5 main classes of heavy chain constant domains. Each class defines the IgM, IgG, IgA, IgD, and IgE isotypes. IgG can be split into 4 subclasses, IgG1, IgG2, IgG3, and IgG4, each with its own biologic properties, and IgA can similarly be split into IgA1 and IgA2.
Copyright 2010 American Academy of Allergy, Asthma & Immunology. Published by Mosby, Inc. All rights reserved.
Figures
References
-
- Williams AF, Barclay AN. The immunoglobulin superfamily--domains for cell surface recognition. Annu. Rev. Immunol. 1988;6:381–405. - PubMed
-
- Harpaz Y, Chothia C. Many of the immunoglobulin superfamily domains in cell adhesion molecules and surface receptors belong to a new structural set which is close to that containing variable domains. J. Mol. Biol. 1994;238:528–539. - PubMed
-
- Torres RM, Imboden J, Schroeder HW., Jr. Antigen receptor genes, gene products, and co-receptors. In: Rich RR, Fleisher TA, Shearer WT, Schroeder HW Jr., Frew AJ, Weyand CM, editors. Clinical Immunology: Principles and Practice. 3 ed. London: Mosby Elsevier; 2008. pp. 53–77.
-
- Hamers-Casterman C, Atarhouch T, Muyldermans S, Robinson G, Hamers C, Songa EB, Bendahman N, Hamers R. Naturally occurring antibodies devoid of light chains. Nature. 1993;363:446–448. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
