

Epithelial interleukin-4 receptor expression promotes colon tumor growth
- PMID: 20176658
- PMCID: PMC2878360
- DOI: 10.1093/carcin/bgq044
Epithelial interleukin-4 receptor expression promotes colon tumor growth
Abstract
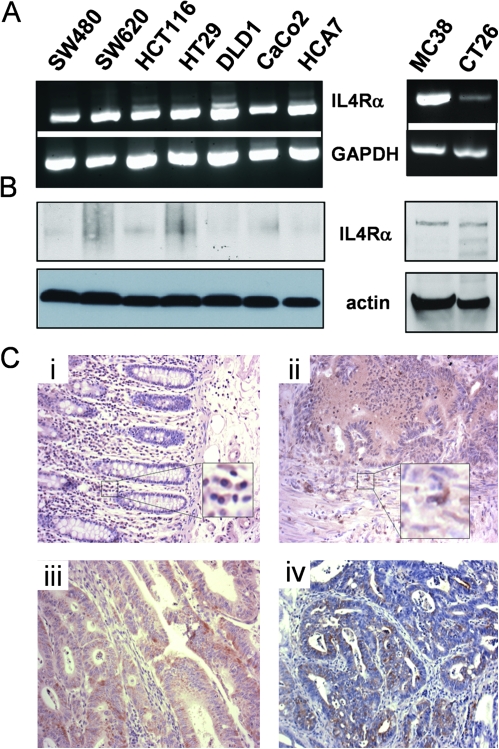
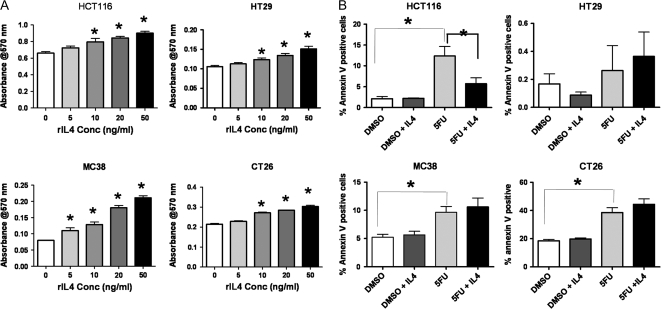
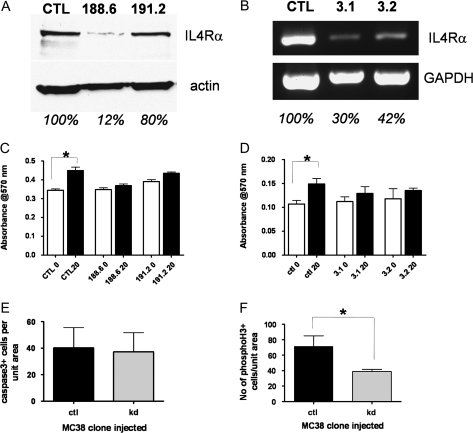
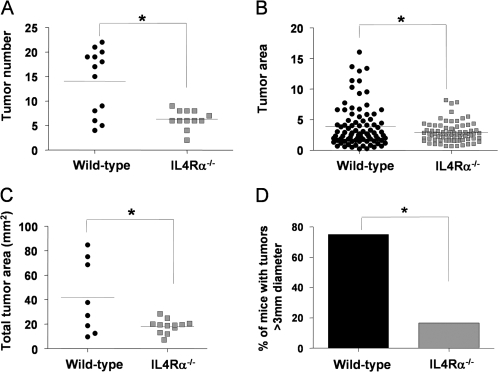
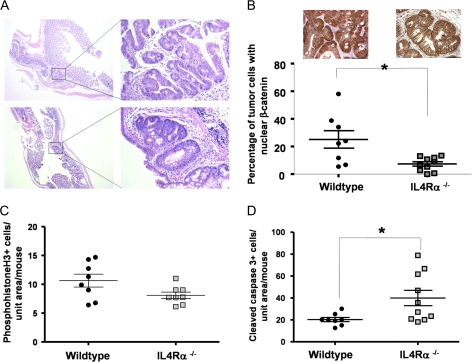
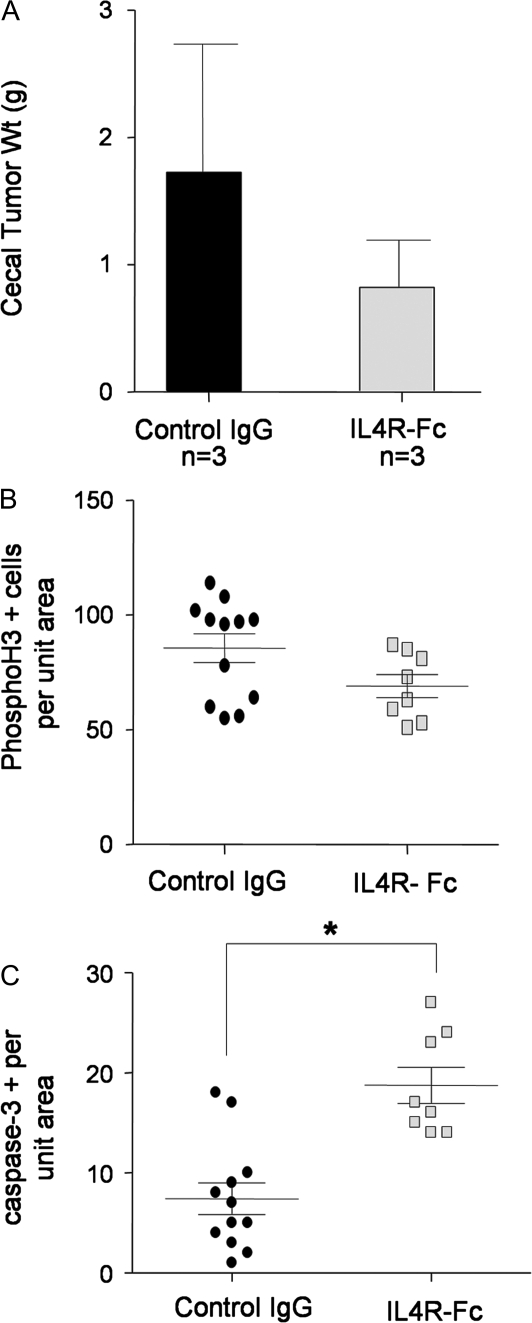
Inflammatory mediators are of considerable interest as potential therapeutic targets in various cancers. Here we investigate whether interleukin (IL)-4 receptor alpha (IL4Ralpha), a component of the receptor complex for the T helper 2 cytokines IL4 and IL13, plays a role in colonic tumorigenesis. IL4Ralpha protein expression was seen in tumor cells of 28/48 human colon adenocarcinomas on a tissue microarray. In human and murine colon tumor cell lines analyzed in vitro, all of which expressed IL4Ralpha, treatment with exogenous ligand resulted in dose-dependent increases in proliferation. IL4 decreased apoptosis only in HCT116 cells. An orthotopic allograft model was used to determine in vivo effects of tumor cell-specific IL4Ra ablation. MC38 murine tumor cells with the IL4Ra gene knocked down showed reduced proliferation but no difference in apoptosis compared with controls after implantation in ceca of syngeneic mice. Mice null for IL4Ra and wild-type controls were treated with azoxymethane and dextran sulfate sodium to induce tumor formation. Mice with global deletion of IL4Ra had significantly fewer and smaller tumors. Reduced tumorigenicity correlated with decreased proliferation and increased apoptosis. Systemic blockade of IL4Ralpha-IL4 interactions with a chimeric soluble receptor protein gave similar results in the cecal implant model. Thus, IL4Ralpha, a component of the IL4R and IL13R, contributes to tumor formation in a mouse model of colitis-associated cancer. Proliferation appears to be directly mediated via IL4Ralpha on the epithelial tumor cells. Survival may be an indirect response mediated via other host cells. Our results support therapeutic targeting of IL4Ralpha in colon cancer.
Figures
References
-
- Mantovani A, et al. Cancer-related inflammation. Nature. 2008;454:436–444. - PubMed
-
- Pohl C, et al. Chronic inflammatory bowel disease and cancer. Hepatogastroenterology. 2000;47:57–70. - PubMed
-
- Itzkowitz SH, et al. Inflammation and cancer IV. Colorectal cancer in inflammatory bowel disease: the role of inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004;287:G7–G17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
