

Depleting syndecan-4+ T lymphocytes using toxin-bearing dendritic cell-associated heparan sulfate proteoglycan-dependent integrin ligand: a new opportunity for treating activated T cell-driven disease
- PMID: 20176742
- PMCID: PMC2963166
- DOI: 10.4049/jimmunol.0903250
Depleting syndecan-4+ T lymphocytes using toxin-bearing dendritic cell-associated heparan sulfate proteoglycan-dependent integrin ligand: a new opportunity for treating activated T cell-driven disease
Abstract
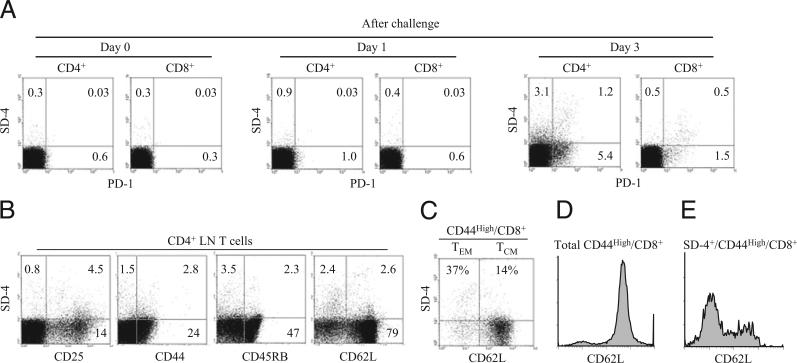
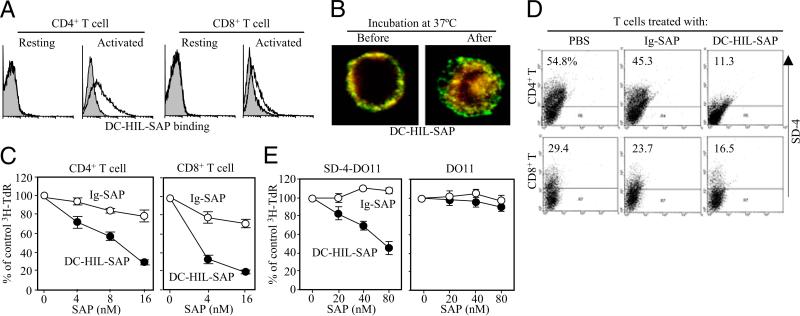
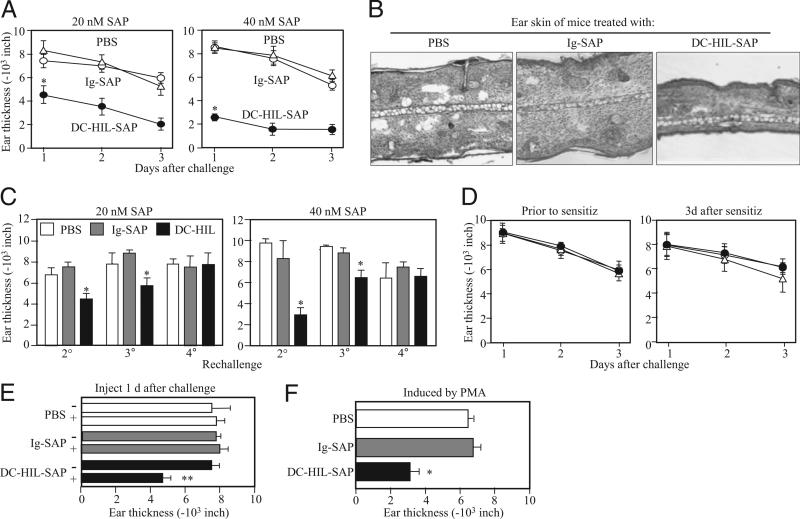
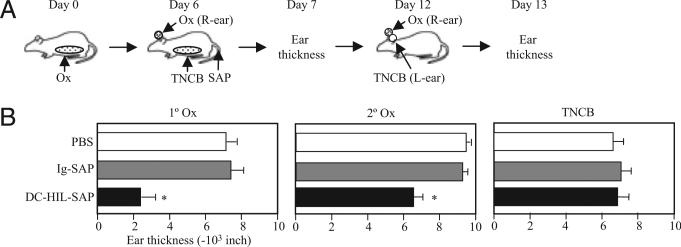
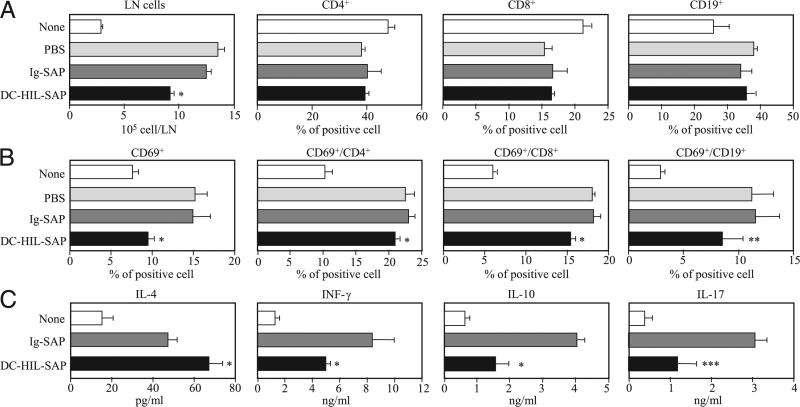
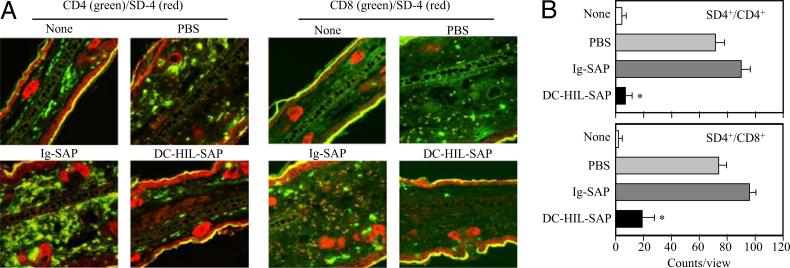
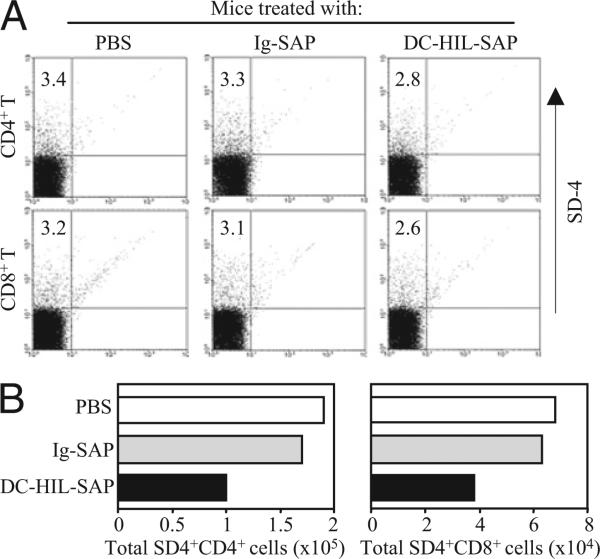
Because syndecan-4 (SD-4) is expressed by some (but not all) T cells following activation and serves as the exclusive ligand of dendritic cell-associated heparan sulfate proteoglycan-dependent integrin ligand (DC-HIL), we envisioned the DC-HIL/SD-4 pathway to be a therapeutic target for conditions mediated by selectively activated T cells. We conjugated soluble DC-HIL receptor with the toxin saporin (SAP; DC-HIL-SAP) and showed it to bind activated (but not resting) T cells and become internalized by and deplete SD-4(+) T cells. In hapten-sensitized mice, DC-HIL-SAP injected i.v. prior to hapten challenge led to markedly suppressed contact hypersensitivity responses that lasted 3 wk and were restricted to the hapten to which the mice were originally sensitized. Such suppression was not observed when DC-HIL-SAP was applied during sensitization. Moreover, the same infusion of DC-HIL-SAP produced almost complete disappearance of SD-4(+) cells in haptenated skin and a 40% reduction of such cells within draining lymph nodes. Our results provide a strong rationale for exploring use of toxin-conjugated DC-HIL to treat activated T cell-driven disease in humans.
Figures
Similar articles
-
Syndecan-4 mediates the coinhibitory function of DC-HIL on T cell activation.J Immunol. 2007 Nov 1;179(9):5778-84. doi: 10.4049/jimmunol.179.9.5778. J Immunol. 2007. PMID: 17947650
-
The DC-HIL/syndecan-4 pathway regulates autoimmune responses through myeloid-derived suppressor cells.J Immunol. 2014 Mar 15;192(6):2576-84. doi: 10.4049/jimmunol.1301857. Epub 2014 Feb 10. J Immunol. 2014. PMID: 24516197 Free PMC article.
-
The DC-HIL/syndecan-4 pathway inhibits human allogeneic T-cell responses.Eur J Immunol. 2009 Apr;39(4):965-74. doi: 10.1002/eji.200838990. Eur J Immunol. 2009. PMID: 19350579 Free PMC article.
-
Sézary syndrome cells overexpress syndecan-4 bearing distinct heparan sulfate moieties that suppress T-cell activation by binding DC-HIL and trapping TGF-beta on the cell surface.Blood. 2011 Mar 24;117(12):3382-90. doi: 10.1182/blood-2010-08-302034. Epub 2011 Jan 20. Blood. 2011. PMID: 21252093 Free PMC article.
-
Immunological mechanisms of contact hypersensitivity in mice.APMIS. 2012 Jan;120(1):1-27. doi: 10.1111/j.1600-0463.2011.02832.x. Epub 2011 Nov 11. APMIS. 2012. PMID: 22151305 Review.
Cited by
-
The DC-HIL ligand syndecan-4 is a negative regulator of T-cell allo-reactivity responsible for graft-versus-host disease.Immunology. 2013 Feb;138(2):173-82. doi: 10.1111/imm.12027. Immunology. 2013. PMID: 23113638 Free PMC article.
-
A translational perspective of the malignant hematopoietic proteoglycome.Cell Biosci. 2025 Feb 20;15(1):25. doi: 10.1186/s13578-025-01360-7. Cell Biosci. 2025. PMID: 39980017 Free PMC article. Review.
-
Critical role for syndecan-4 in dendritic cell migration during development of allergic airway inflammation.Nat Commun. 2015 Jul 13;6:7554. doi: 10.1038/ncomms8554. Nat Commun. 2015. PMID: 26165408
-
Streptavidin-Saporin: Converting Biotinylated Materials into Targeted Toxins.Toxins (Basel). 2023 Feb 27;15(3):181. doi: 10.3390/toxins15030181. Toxins (Basel). 2023. PMID: 36977072 Free PMC article. Review.
-
Glycoprotein nonmetastatic melanoma protein B: A key mediator and an emerging therapeutic target in autoimmune diseases.FASEB J. 2020 Jul;34(7):8810-8823. doi: 10.1096/fj.202000651. Epub 2020 May 23. FASEB J. 2020. PMID: 32445534 Free PMC article. Review.
References
-
- Chambers CA, Allison JP. Co-stimulation in T cell responses. Curr. Opin. Immunol. 1997;9:396–404. - PubMed
-
- Carreno BM, Collins M. The B7 family of ligands and its receptors: new pathways for costimulation and inhibition of immune responses. Annu. Rev. Immunol. 2002;20:29–53. - PubMed
-
- Carreno BM, Bennett F, Chau TA, Ling V, Luxenberg D, Jussif J, Baroja ML, Madrenas J. CTLA-4 (CD152) can inhibit T cell activation by two different mechanisms depending on its level of cell surface expression. J. Immunol. 2000;165:1352–1356. - PubMed
-
- Tivol EA, Borriello F, Schweitzer AN, Lynch WP, Bluestone JA, Sharpe AH. Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4. Immunity. 1995;3:541–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
