

The Rickettsia conorii autotransporter protein Sca1 promotes adherence to nonphagocytic mammalian cells
- PMID: 20176791
- PMCID: PMC2863548
- DOI: 10.1128/IAI.01165-09
The Rickettsia conorii autotransporter protein Sca1 promotes adherence to nonphagocytic mammalian cells
Abstract
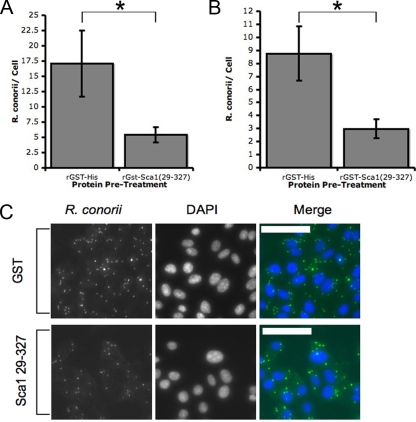
The pathogenesis of spotted fever group (SFG) Rickettsia species, including R. conorii and R. rickettsii, is acutely dependent on adherence to and invasion of host cells, including cells of the mammalian endothelial system. Bioinformatic analyses of several rickettsia genomes revealed the presence of a cohort of genes designated sca genes that are predicted to encode proteins with homology to autotransporter proteins of Gram-negative bacteria. Previous work demonstrated that three members of this family, rOmpA (Sca0), Sca2, and rOmpB (Sca5) are involved in the interaction with mammalian cells; however, very little was known about the function of other conserved rickettsial Sca proteins. Here we demonstrate that sca1, a gene present in nearly all SFG rickettsia genomes, is actively transcribed and expressed in R. conorii cells. Alignment of Sca1 sequences from geographically diverse SFG Rickettsia species showed that there are high degrees of sequence identity and conservation of these sequences, suggesting that Sca1 may have a conserved function. Using a heterologous expression system, we demonstrated that production of R. conorii Sca1 in the Escherichia coli outer membrane is sufficient to mediate attachment to but not invasion of a panel of cultured mammalian epithelial and endothelial cells. Furthermore, preincubation of a recombinant Sca1 peptide with host cells blocked R. conorii cell association. Together, these results demonstrate that attachment to mammalian cells can be uncoupled from the entry process and that Sca1 is involved in the adherence of R. conorii to host cells.
Figures





Similar articles
-
OmpA-mediated rickettsial adherence to and invasion of human endothelial cells is dependent upon interaction with α2β1 integrin.Cell Microbiol. 2013 May;15(5):727-41. doi: 10.1111/cmi.12068. Epub 2012 Dec 6. Cell Microbiol. 2013. PMID: 23145974 Free PMC article.
-
The Sca2 autotransporter protein from Rickettsia conorii is sufficient to mediate adherence to and invasion of cultured mammalian cells.Infect Immun. 2009 Dec;77(12):5272-80. doi: 10.1128/IAI.00201-09. Epub 2009 Oct 5. Infect Immun. 2009. PMID: 19805531 Free PMC article.
-
Identification and characterization of the mammalian association and actin-nucleating domains in the Rickettsia conorii autotransporter protein, Sca2.Cell Microbiol. 2012 Sep;14(9):1485-95. doi: 10.1111/j.1462-5822.2012.01815.x. Epub 2012 Jun 5. Cell Microbiol. 2012. PMID: 22612237 Free PMC article.
-
[Emerging rickettsioses].Parassitologia. 2004 Jun;46(1-2):123-6. Parassitologia. 2004. PMID: 15305700 Review. Italian.
-
Progress in rickettsial genome analysis from pioneering of Rickettsia prowazekii to the recent Rickettsia typhi.Ann N Y Acad Sci. 2005 Dec;1063:13-25. doi: 10.1196/annals.1355.003. Ann N Y Acad Sci. 2005. PMID: 16481486 Review.
Cited by
-
OmpA-mediated rickettsial adherence to and invasion of human endothelial cells is dependent upon interaction with α2β1 integrin.Cell Microbiol. 2013 May;15(5):727-41. doi: 10.1111/cmi.12068. Epub 2012 Dec 6. Cell Microbiol. 2013. PMID: 23145974 Free PMC article.
-
The Orientia tsutsugamushi ScaB Autotransporter Protein Is Required for Adhesion and Invasion of Mammalian Cells.Front Microbiol. 2021 Feb 4;12:626298. doi: 10.3389/fmicb.2021.626298. eCollection 2021. Front Microbiol. 2021. PMID: 33613493 Free PMC article.
-
Cells within cells: Rickettsiales and the obligate intracellular bacterial lifestyle.Nat Rev Microbiol. 2021 Jun;19(6):375-390. doi: 10.1038/s41579-020-00507-2. Epub 2021 Feb 9. Nat Rev Microbiol. 2021. PMID: 33564174 Review.
-
Identification of an autotransporter peptidase of Rickettsia rickettsii responsible for maturation of surface exposed autotransporters.PLoS Pathog. 2023 Jul 31;19(7):e1011527. doi: 10.1371/journal.ppat.1011527. eCollection 2023 Jul. PLoS Pathog. 2023. PMID: 37523399 Free PMC article.
-
Metagenome diversity illuminates origins of pathogen effectors.bioRxiv [Preprint]. 2023 Feb 27:2023.02.26.530123. doi: 10.1101/2023.02.26.530123. bioRxiv. 2023. Update in: mBio. 2024 May 8;15(5):e0075923. doi: 10.1128/mbio.00759-23. PMID: 36909625 Free PMC article. Updated. Preprint.
References
-
- Billings, A. N., H. M. Feng, J. P. Olano, and D. H. Walker. 2001. Rickettsial infection in murine models activates an early anti-rickettsial effect mediated by NK cells and associated with production of gamma interferon. Am. J. Trop. Med. Hyg. 65:52-56. - PubMed
-
- Blanc, G., M. Ngwamidiba, H. Ogata, P. E. Fournier, J. M. Claverie, and D. Raoult. 2005. Molecular evolution of rickettsia surface antigens: evidence of positive selection. Mol. Biol. Evol. 22:2073-2083. - PubMed
-
- Braun, L., H. Ohayon, and P. Cossart. 1998. The InIB protein of Listeria monocytogenes is sufficient to promote entry into mammalian cells. Mol. Microbiol. 27:1077-1087. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
