

Diffusion of a soluble protein, photoactivatable GFP, through a sensory cilium
- PMID: 20176852
- PMCID: PMC2828910
- DOI: 10.1085/jgp.200910322
Diffusion of a soluble protein, photoactivatable GFP, through a sensory cilium
Abstract
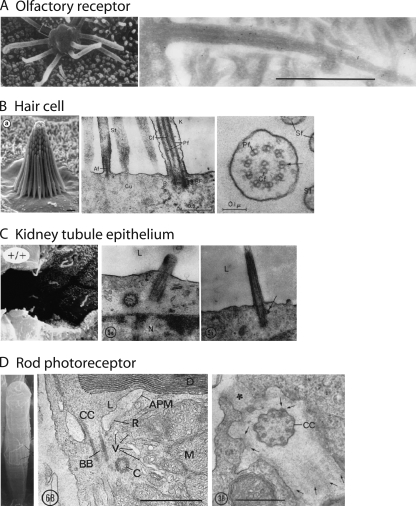
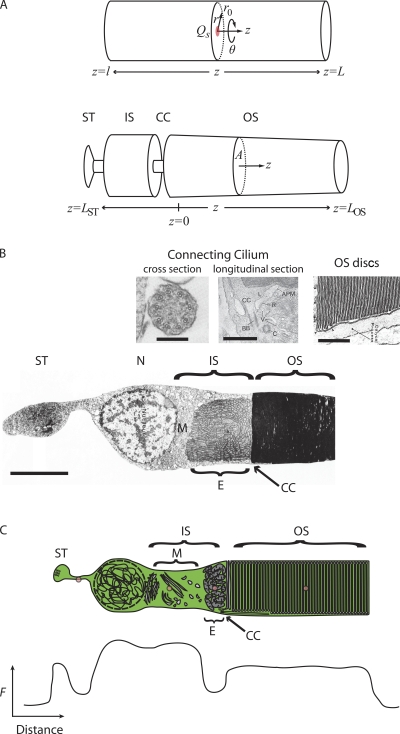
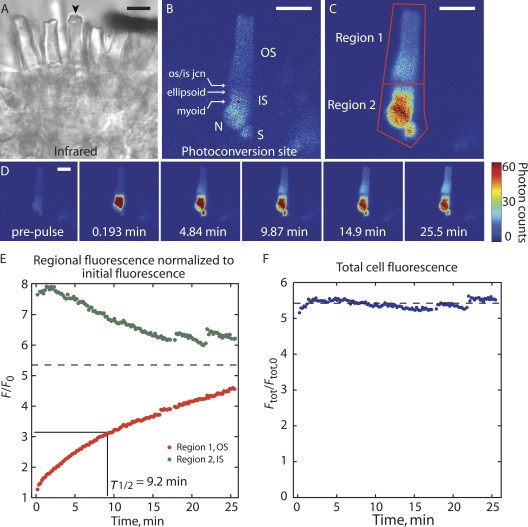
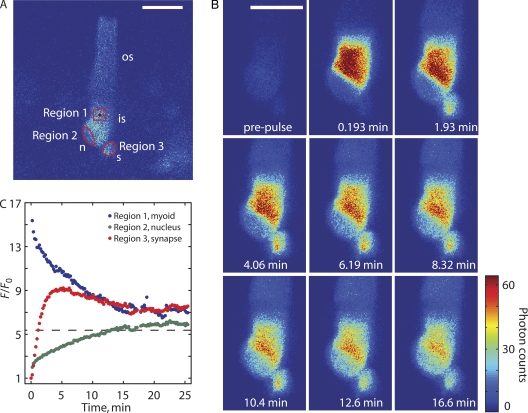
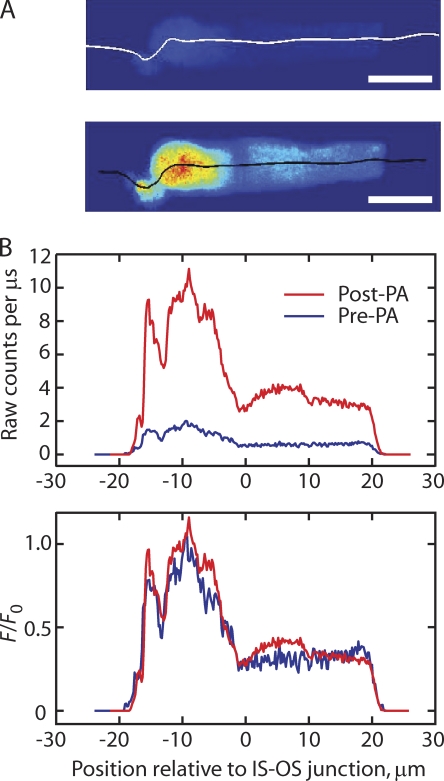
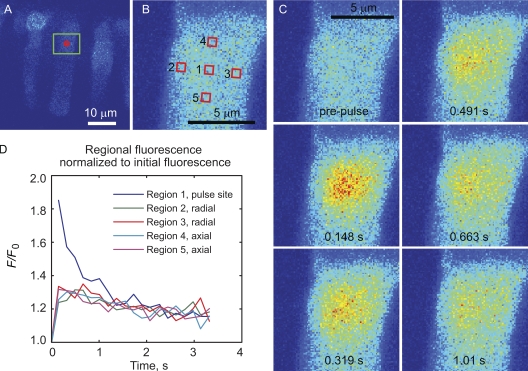
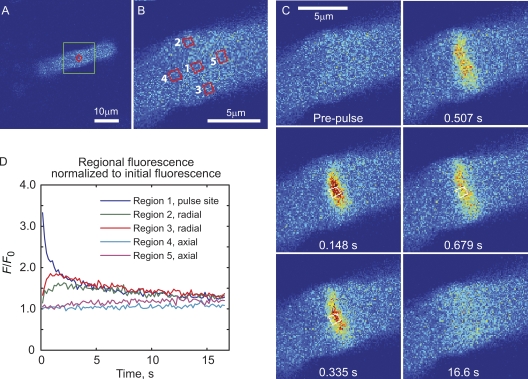
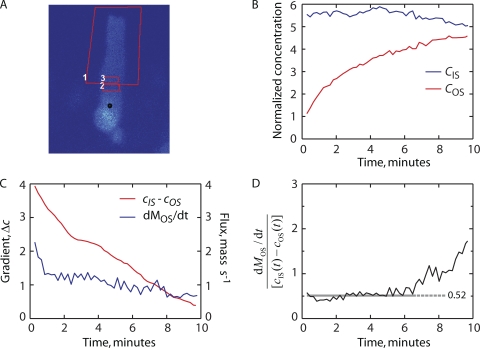
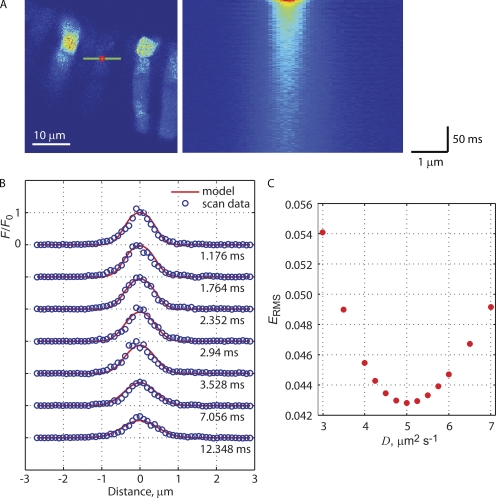
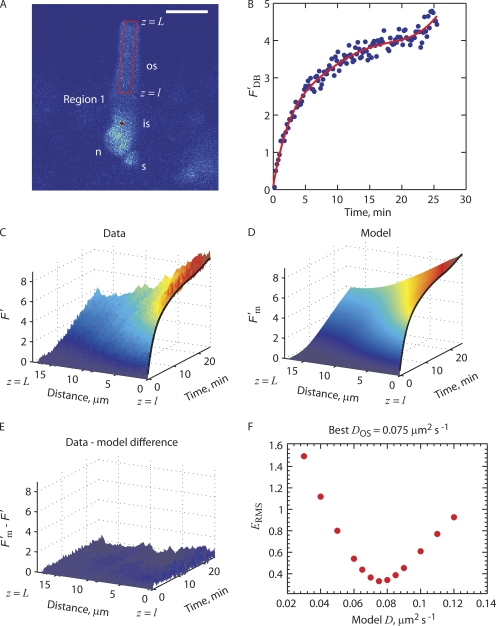
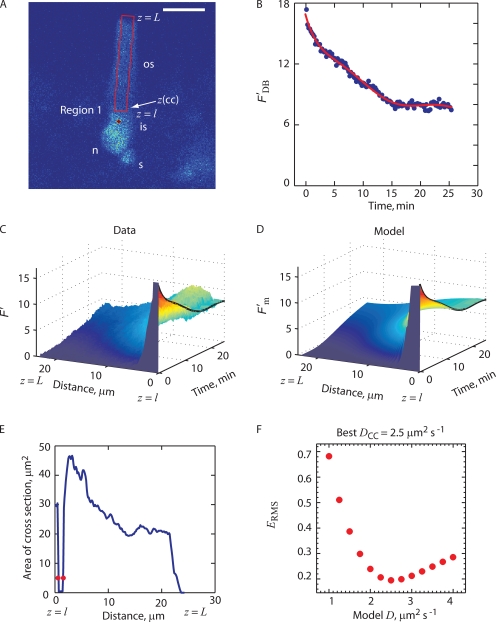
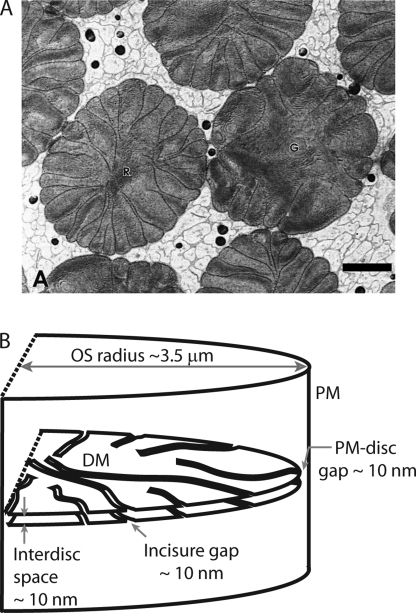
Transport of proteins to and from cilia is crucial for normal cell function and survival, and interruption of transport has been implicated in degenerative and neoplastic diseases. It has been hypothesized that the ciliary axoneme and structures adjacent to and including the basal bodies of cilia impose selective barriers to the movement of proteins into and out of the cilium. To examine this hypothesis, using confocal and multiphoton microscopy we determined the mobility of the highly soluble photoactivatable green fluorescent protein (PAGFP) in the connecting cilium (CC) of live Xenopus retinal rod photoreceptors, and in the contiguous subcellular compartments bridged by the CC, the inner segment (IS) and the outer segment (OS). The estimated axial diffusion coefficients are D(CC) = 2.8 +/- 0.3, D(IS) = 5.2 +/- 0.6, and D(OS) = 0.079 +/- 0.009 microm(2) s(-1). The results establish that the CC does not pose a major barrier to protein diffusion within the rod cell. However, the results also reveal that axial diffusion in each of the rod's compartments is substantially retarded relative to aqueous solution: the axial diffusion of PAGFP was retarded approximately 18-, 32- and 1,000-fold in the IS, CC, and OS, respectively, with approximately 20-fold of the reduction in the OS attributable to tortuosity imposed by the lamellar disc membranes. Previous investigation of PAGFP diffusion in passed, spherical Chinese hamster ovary cells yielded D(CHO) = 20 microm(2) s(-1), and estimating cytoplasmic viscosity as D(aq)/D(CHO) = 4.5, the residual 3- to 10-fold reduction in PAGFP diffusion is ascribed to sub-optical resolution structures in the IS, CC, and OS compartments.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
