

Characterization of plant sulfiredoxin and role of sulphinic form of 2-Cys peroxiredoxin
- PMID: 20176891
- PMCID: PMC2837264
- DOI: 10.1093/jxb/erq016
Characterization of plant sulfiredoxin and role of sulphinic form of 2-Cys peroxiredoxin
Abstract
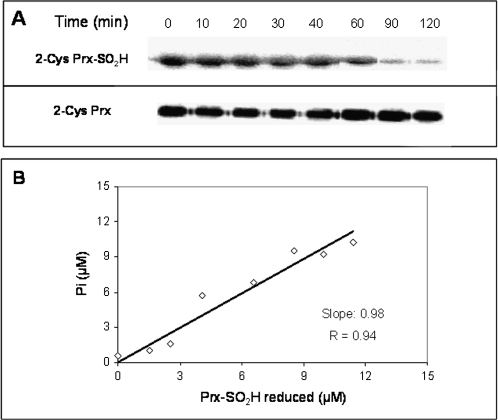
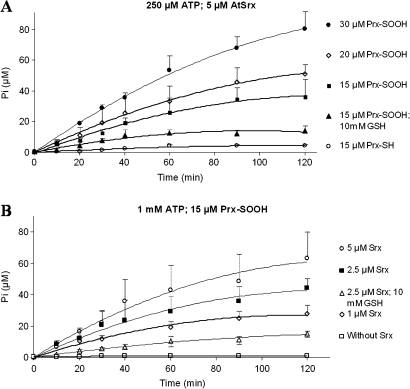
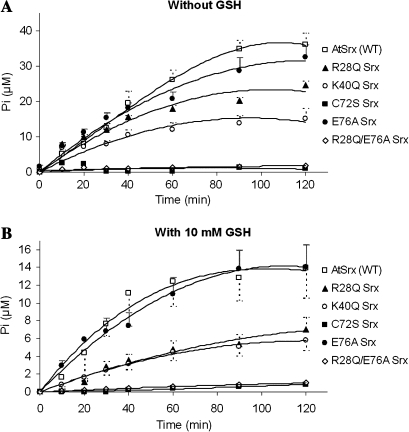
The antioxidant function of 2-Cys peroxiredoxin (Prx) involves the oxidation of its conserved peroxidatic cysteine to sulphenic acid that is recycled by a reductor agent. In conditions of oxidative stress, the peroxidatic cysteine can be overoxidized to sulphinic acid inactivating the Prx. An enzyme recently discovered, named sulfiredoxin (Srx), reduces the sulphinic 2-Cys Prx (Prx-SO(2)H). To explore the physiological functions of Srx in plants we have cloned, expressed and purified to homogeneity a Srx from Arabidopsis thaliana (AtSrx), as well as five variants by site-directed mutagenesis on amino acids involved in its activity. The activity of sulfiredoxin, determined by a new method, is dependent on the concentration of the sulphinic form of Prx and the conserved Srx is capable of regenerating the functionality of both pea and Arabidopsis Prx-SO(2)H. Molecular modelling of AtSrx and the facts that the R28Q variant shows a partial inactivation, that the activity of the E76A variant is equivalent to that of the native enzyme and that the double mutation R28Q/E76A abolishes the enzymatic activity suggests that the pair His100-Glu76 may be involved in the activation of C72 in the absence of R28. The knock-out mutant plants without Srx or 2-Cys Prx exhibited phenotypical differences under growth conditions of 16 h light, probably due to the signalling role of the sulphinic form of Prx. These mutants showed more susceptibility to oxidative stress than wild-type plants. This work presents the first systematic biochemical characterization of the Srx/Prx system from plants and contributes to a better understanding of its physiological function.
Figures






Similar articles
-
The contribution of NADPH thioredoxin reductase C (NTRC) and sulfiredoxin to 2-Cys peroxiredoxin overoxidation in Arabidopsis thaliana chloroplasts.J Exp Bot. 2015 May;66(10):2957-66. doi: 10.1093/jxb/eru512. Epub 2015 Jan 5. J Exp Bot. 2015. PMID: 25560178 Free PMC article.
-
The dual-targeted plant sulfiredoxin retroreduces the sulfinic form of atypical mitochondrial peroxiredoxin.Plant Physiol. 2011 Feb;155(2):944-55. doi: 10.1104/pp.110.166504. Epub 2010 Dec 7. Plant Physiol. 2011. PMID: 21139087 Free PMC article.
-
The Arabidopsis thaliana sulfiredoxin is a plastidic cysteine-sulfinic acid reductase involved in the photooxidative stress response.Plant J. 2007 Feb;49(3):505-14. doi: 10.1111/j.1365-313X.2006.02969.x. Epub 2007 Jan 1. Plant J. 2007. PMID: 17217469
-
The sulfiredoxin-peroxiredoxin (Srx-Prx) axis in cell signal transduction and cancer development.Cancer Lett. 2015 Oct 1;366(2):150-9. doi: 10.1016/j.canlet.2015.07.002. Epub 2015 Jul 10. Cancer Lett. 2015. PMID: 26170166 Free PMC article. Review.
-
Sulfiredoxin, the cysteine sulfinic acid reductase specific to 2-Cys peroxiredoxin: its discovery, mechanism of action, and biological significance.Kidney Int Suppl. 2007 Aug;(106):S3-8. doi: 10.1038/sj.ki.5002380. Kidney Int Suppl. 2007. PMID: 17653208 Review.
Cited by
-
Thioredoxin Network in Plant Mitochondria: Cysteine S-Posttranslational Modifications and Stress Conditions.Front Plant Sci. 2020 Sep 23;11:571288. doi: 10.3389/fpls.2020.571288. eCollection 2020. Front Plant Sci. 2020. PMID: 33072147 Free PMC article. Review.
-
Disruption of both chloroplastic and cytosolic FBPase genes results in a dwarf phenotype and important starch and metabolite changes in Arabidopsis thaliana.J Exp Bot. 2015 May;66(9):2673-89. doi: 10.1093/jxb/erv062. Epub 2015 Mar 5. J Exp Bot. 2015. PMID: 25743161 Free PMC article.
-
The contribution of NADPH thioredoxin reductase C (NTRC) and sulfiredoxin to 2-Cys peroxiredoxin overoxidation in Arabidopsis thaliana chloroplasts.J Exp Bot. 2015 May;66(10):2957-66. doi: 10.1093/jxb/eru512. Epub 2015 Jan 5. J Exp Bot. 2015. PMID: 25560178 Free PMC article.
-
Overoxidation of 2-Cys peroxiredoxin in prokaryotes: cyanobacterial 2-Cys peroxiredoxins sensitive to oxidative stress.J Biol Chem. 2010 Nov 5;285(45):34485-92. doi: 10.1074/jbc.M110.160465. Epub 2010 Aug 24. J Biol Chem. 2010. PMID: 20736168 Free PMC article.
-
Evidence that glutathione and the glutathione system efficiently recycle 1-cys sulfiredoxin in vivo.Antioxid Redox Signal. 2015 Mar 20;22(9):731-43. doi: 10.1089/ars.2014.5998. Epub 2015 Jan 8. Antioxid Redox Signal. 2015. PMID: 25387359 Free PMC article.
References
-
- Baginski ES, Epstein E, Zak B. Review of phosphate methodologies. Annals of Clinical and Laboratory Science. 1975;5:399–416. - PubMed
-
- Barranco-Medina S, Kakorin S, Lázaro JJ, Dietz KJ. Thermodynamics of the dimmer-decamer transition of reduced human and plant 2-Cys peroxiredoxin. Biochemistry. 2008;47:7196–7204. - PubMed
-
- Bernier-Villamor L, Navarro E, Sevilla F, Lázaro JJ. Cloning and characterization of a 2-Cys peroxiredoxin from Pisum sativum. Journal of Experimental Botany. 2004;55:2191–2199. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
