

Automated microscopy for high-content RNAi screening
- PMID: 20176920
- PMCID: PMC2828931
- DOI: 10.1083/jcb.200910105
Automated microscopy for high-content RNAi screening
Abstract
Fluorescence microscopy is one of the most powerful tools to investigate complex cellular processes such as cell division, cell motility, or intracellular trafficking. The availability of RNA interference (RNAi) technology and automated microscopy has opened the possibility to perform cellular imaging in functional genomics and other large-scale applications. Although imaging often dramatically increases the content of a screening assay, it poses new challenges to achieve accurate quantitative annotation and therefore needs to be carefully adjusted to the specific needs of individual screening applications. In this review, we discuss principles of assay design, large-scale RNAi, microscope automation, and computational data analysis. We highlight strategies for imaging-based RNAi screening adapted to different library and assay designs.
Figures
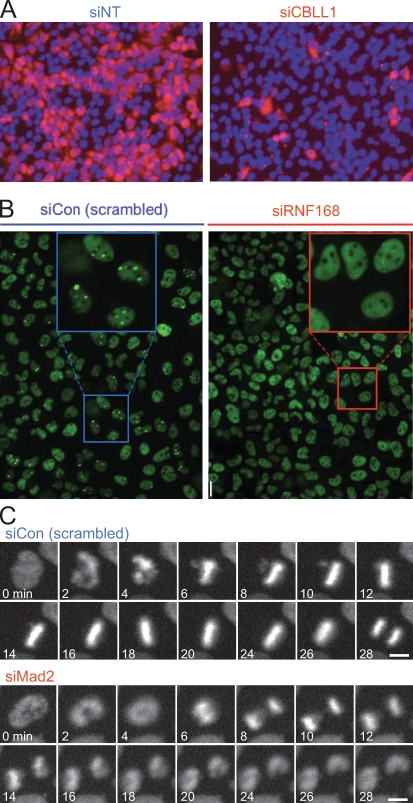
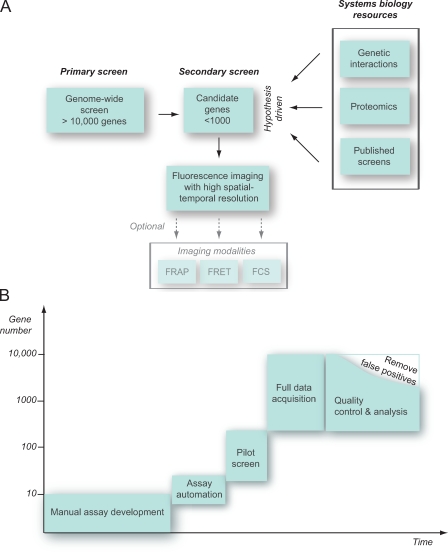
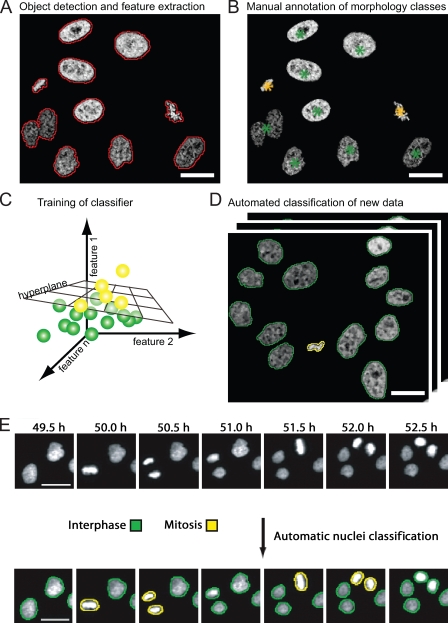
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
