

Condensin complexes regulate mitotic progression and interphase chromatin structure in embryonic stem cells
- PMID: 20176923
- PMCID: PMC2828918
- DOI: 10.1083/jcb.200908026
Condensin complexes regulate mitotic progression and interphase chromatin structure in embryonic stem cells
Abstract
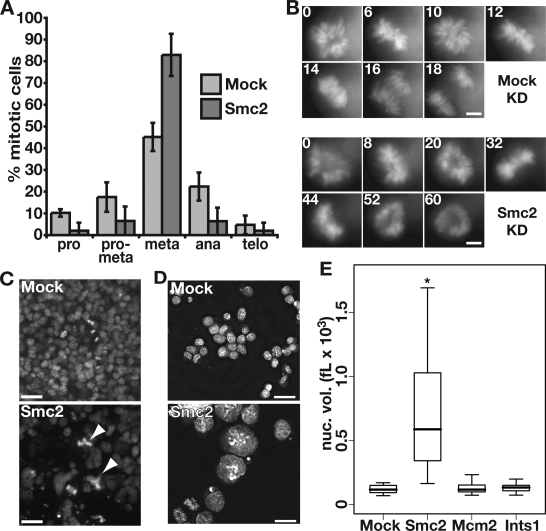
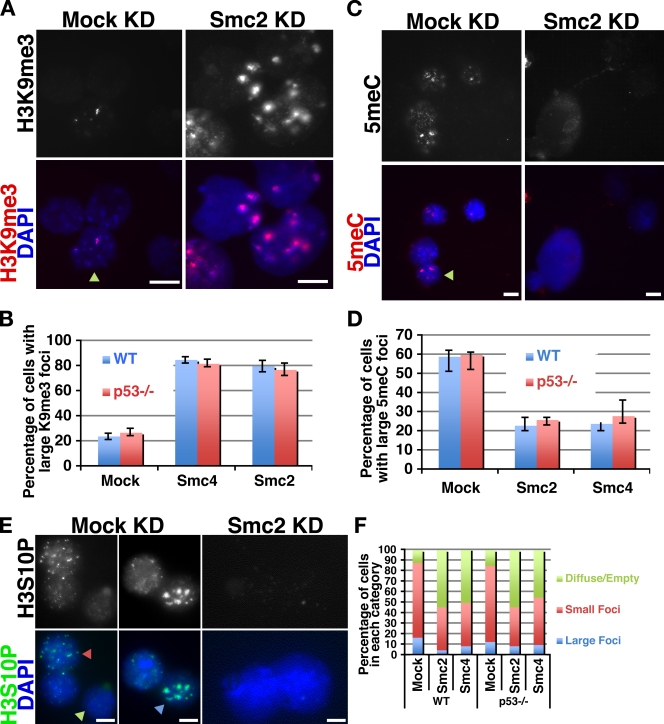
In an RNA interference screen interrogating regulators of mouse embryonic stem (ES) cell chromatin structure, we previously identified 62 genes required for ES cell viability. Among these 62 genes were Smc2 and -4, which are core components of the two mammalian condensin complexes. In this study, we show that for Smc2 and -4, as well as an additional 49 of the 62 genes, knockdown (KD) in somatic cells had minimal effects on proliferation or viability. Upon KD, Smc2 and -4 exhibited two phenotypes that were unique to ES cells and unique among the ES cell-lethal targets: metaphase arrest and greatly enlarged interphase nuclei. Nuclear enlargement in condensin KD ES cells was caused by a defect in chromatin compaction rather than changes in DNA content. The altered compaction coincided with alterations in the abundance of several epigenetic modifications. These data reveal a unique role for condensin complexes in interphase chromatin compaction in ES cells.
Figures




Similar articles
-
Fission yeast condensin contributes to interphase chromatin organization and prevents transcription-coupled DNA damage.Genome Biol. 2020 Nov 5;21(1):272. doi: 10.1186/s13059-020-02183-0. Genome Biol. 2020. PMID: 33153481 Free PMC article.
-
Mitotic post-translational modifications of histones promote chromatin compaction in vitro.Open Biol. 2017 Sep;7(9):170076. doi: 10.1098/rsob.170076. Open Biol. 2017. PMID: 28903997 Free PMC article.
-
Disturbance in function and expression of condensin affects chromosome compaction in HeLa cells.Cell Biol Int. 2011 Jul;35(7):735-40. doi: 10.1042/CBI20100646. Cell Biol Int. 2011. PMID: 21395557
-
Condensin Regulation of Genome Architecture.J Cell Physiol. 2017 Jul;232(7):1617-1625. doi: 10.1002/jcp.25702. Epub 2017 Jan 5. J Cell Physiol. 2017. PMID: 27888504 Review.
-
Three-step model for condensin activation during mitotic chromosome condensation.Cell Cycle. 2010 Aug 15;9(16):3243-55. doi: 10.4161/cc.9.16.12620. Epub 2010 Aug 7. Cell Cycle. 2010. PMID: 20703077 Review.
Cited by
-
Histone Code and Higher-Order Chromatin Folding: A Hypothesis.Genom Comput Biol. 2017;3(2):e41. doi: 10.18547/gcb.2017.vol3.iss2.e41. Epub 2017 Jan 30. Genom Comput Biol. 2017. PMID: 31245531 Free PMC article.
-
Adaptation of the AID system for stem cell and transgenic mouse research.Stem Cell Res. 2020 Dec;49:102078. doi: 10.1016/j.scr.2020.102078. Epub 2020 Nov 5. Stem Cell Res. 2020. PMID: 33202307 Free PMC article.
-
Single-cell atlas of the small intestine throughout the human lifespan demonstrates unique features of fetal immune cells.Mucosal Immunol. 2024 Aug;17(4):599-617. doi: 10.1016/j.mucimm.2024.03.011. Epub 2024 Mar 28. Mucosal Immunol. 2024. PMID: 38555026 Free PMC article.
-
Fission yeast condensin contributes to interphase chromatin organization and prevents transcription-coupled DNA damage.Genome Biol. 2020 Nov 5;21(1):272. doi: 10.1186/s13059-020-02183-0. Genome Biol. 2020. PMID: 33153481 Free PMC article.
-
Open chromatin in pluripotency and reprogramming.Nat Rev Mol Cell Biol. 2011 Jan;12(1):36-47. doi: 10.1038/nrm3036. Nat Rev Mol Cell Biol. 2011. PMID: 21179060 Free PMC article. Review.
