

Arabidopsis thaliana life without phytochromes
- PMID: 20176939
- PMCID: PMC2842051
- DOI: 10.1073/pnas.0910446107
Arabidopsis thaliana life without phytochromes
Abstract
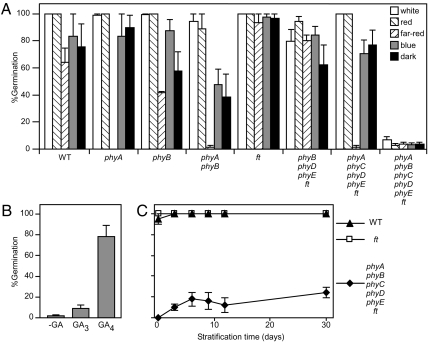
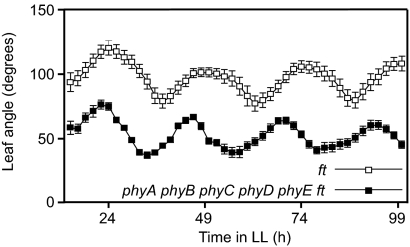
Plants use light as a source of energy for photosynthesis and as a source of environmental information perceived by photoreceptors. Testing whether plants can complete their cycle if light provides energy but no information about the environment requires a plant devoid of phytochromes because all photosynthetically active wavelengths activate phytochromes. Producing such a quintuple mutant of Arabidopsis thaliana has been challenging, but we were able to obtain it in the flowering locus T (ft) mutant background. The quintuple phytochrome mutant does not germinate in the FT background, but it germinates to some extent in the ft background. If germination problems are bypassed by the addition of gibberellins, the seedlings of the quintuple phytochrome mutant exposed to red light produce chlorophyll, indicating that phytochromes are not the sole red-light photoreceptors, but they become developmentally arrested shortly after the cotyledon stage. Blue light bypasses this blockage, rejecting the long-standing idea that the blue-light receptors cryptochromes cannot operate without phytochromes. After growth under white light, returning the quintuple phytochrome mutant to red light resulted in rapid senescence of already expanded leaves and severely impaired expansion of new leaves. We conclude that Arabidopsis development is stalled at several points in the presence of light suitable for photosynthesis but providing no photomorphogenic signal.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
