

Role of alkaline phosphatase from Manduca sexta in the mechanism of action of Bacillus thuringiensis Cry1Ab toxin
- PMID: 20177063
- PMCID: PMC2857145
- DOI: 10.1074/jbc.M109.085266
Role of alkaline phosphatase from Manduca sexta in the mechanism of action of Bacillus thuringiensis Cry1Ab toxin
Abstract
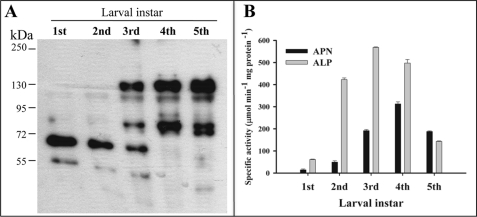
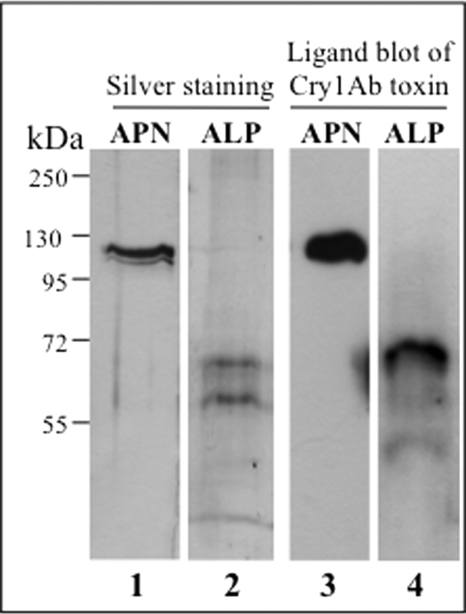
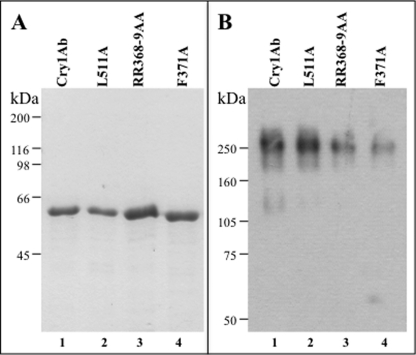
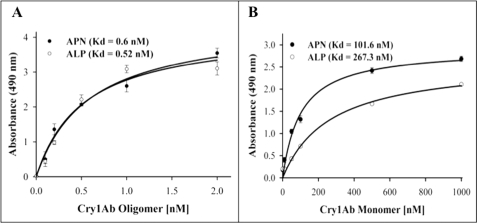
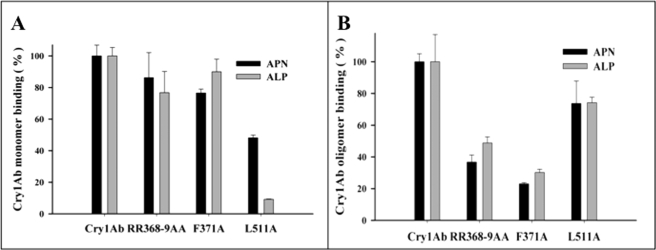
Cry toxins produced by Bacillus thuringiensis have been recognized as pore-forming toxins whose primary action is to lyse midgut epithelial cells in their target insect. In the case of the Cry1A toxins, a prepore oligomeric intermediate is formed after interaction with cadherin receptor. The Cry1A oligomer then interacts with glycosylphosphatidylinositol-anchored receptors. Two Manduca sexta glycosylphosphatidylinositol-anchored proteins, aminopeptidase (APN) and alkaline phosphatase (ALP), have been shown to bind Cry1Ab, although their role in toxicity remains to be determined. Detection of Cry1Ab binding proteins by ligand blot assay revealed that ALP is preferentially expressed earlier during insect development, because it was found in the first larval instars, whereas APN is induced later after the third larval instar. The binding of Cry1Ab oligomer to pure preparations of APN and ALP showed that this toxin structure interacts with both receptors with high affinity (apparent K(d) = 0.6 nM), whereas the monomer showed weaker binding (apparent K(d) = 101.6 and 267.3 nM for APN and ALP, respectively). Several Cry1Ab nontoxic mutants located in the exposed loop 2 of domain II or in beta-16 of domain III were affected in binding to APN and ALP, depending on their oligomeric state. In particular monomers of the nontoxic domain III, the L511A mutant did not bind ALP but retained APN binding, suggesting that initial interaction with ALP is critical for toxicity. Our data suggest that APN and ALP fulfill two roles. First APN and ALP are initial receptors promoting the localization of toxin monomers in the midgut microvilli before interaction with cadherin. Then APN and ALP function as secondary receptors mediating oligomer insertion into the membrane. However, the expression pattern of these receptors and the phenotype of L511A mutant suggest that ALP may have a predominant role in toxin action because Cry toxins are highly effective against the neonate larvae that is the target for pest control programs.
Figures
References
-
- Guo S., Ye S., Liu Y., Wei L., Xue J., Wu H., Song F., Zhang J., Wu X., Huang D., Rao Z. (2009) J. Struct. Biol. 168, 259–266 - PubMed
-
- de Maagd R. A., Bravo A., Berry C., Crickmore N., Schnepf H. E. (2003) Annu. Rev. Genet. 37, 409–433 - PubMed
-
- Jiménez-Juárez N., Muñoz-Garay C., Gómez I., Saab-Rincon G., Damian-Almazo J. Y., Gill S. S., Soberón M., Bravo A. (2007) J. Biol. Chem. 282, 21222–21229 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
